Жаропонижающие средства для детей назначаются педиатром. Но бывают ситуации неотложной помощи при лихорадке, когда ребенку нужно дать лекарство немедленно. Тогда родители берут на себя ответственность и применяют жаропонижающие препараты. Что разрешено давать детям грудного возраста? Чем можно сбить температуру у детей постарше? Какие лекарства самые безопасные?
Гаструляция
Сущность стадии гаструляции заключается в том, что однослойный зародыш - бластула - превращается в многослойный - двух- или трехслойный, называемый гаструлой.
У примитивных хордовых, например у ланцетника, однородная однослойная бластодерма во время гаструляции преобразуется в наружный зародышевый листок - эктодерму - и внутренний зародышевый листок - энтодерму. Энтодерма формирует первичную кишку с полостью внутри - гастроцель. Отверстие, ведущее в гастроцель, называют бластопором или первичным ртом.
У позвоночных образуется еще третий зародышевый листок - мезодерма, занимающая место между экто- и энтодермой. Развитие среднего зародышевого листка, является эволюционным усложнением фазы гаструляции у позвоночных. У более примитивных хордовых животных, таких, как ланцетник, мезодерма обычно образуется в начале следующей после гаструляции фазы - органогенезе.
Процесс гаструляции характеризуется важными, клеточными преобразованиями, такими, как направленные перемещения групп и отдельных клеток, избирательное размножение и сортировка клеток, начало цитодифференцировки и индукционных взаимодействий.
Способы гаструляции различны. Выделяют четыре разновидности, приводящих к преобразованию зародыша из однослойного в многослойный.
Инвагинация - впячивание одного из участков бластодермы внутрь целым пластом. У ланцетника впячиваются клетки вегетативного полюса, у земноводных инвагинация происходит на границе между анимальным и вегетативным полюсами в области серого серпа. Процесс инвагинации возможен только в яйцах с небольшим или средним количеством желтка.
Эпиболия - обрастание мелкими клетками анимального полюса более крупных, отстающих в скорости деления и менее подвижных клеток вегетативного полюса. Такой процесс ярко выражен у земноводных.
Деноминация -расслоение клеток бластодермы на два слоя, лежащих друг над другом. Деламинацию можно наблюдать в дискобластуле зародышей с частичным типом дробления, таких, как пресмыкающиеся, птицы, яйцекладущие млекопитающие. Деламинация проявляется в эмбриобласте плацентарных млекопитающих, приводя к образованию гипобласта и эпибласта.
Иммиграция - перемещение групп или отдельных клеток, не объединенных в единый пласт. Иммиграция встречается у всех зародышей, но в наибольшей степени характерна для второй фазы гаструляции высших позвоночных.
Морфология гаструляции. Инвагинация начинается на вегетативном полюсе. Из-за более быстрого деления клетки анимального полюса разрастаются и толкают внутрь бластулы клетки вегетативного полюса. Вследствие инвагинации бластоцель уменьшается, а гастроцель увеличивается. Одновременно с исчезновением бластоцеля эктодерма и энтодерма приходят в тесный контакт. У ланцетника, как и у всех вторичноротых животных область бластопора превращается в хвостовую часть организма в отличие от первичноротых, у которых бластопор соответствует головной части. Ротовое отверстие у вторичноротых образуется на противоположном бластопору конце зародыша.
Гаструляция у земноводных имеет много общего с гаструляцией ланцетника, но так как в яйцеклетках у них желтка намного больше и расположен он преимущественно на вегетативном полюсе, крупные бластомеры амфибластулы не способны впячиваться внутрь. Инвагинация проходит иначе. На границе между анимальным и вегетативным полюсами в области серого серпа клетки сначала сильно вытягиваются внутрь, принимая вид «колбовидных», а затем тянут за собой клетки поверхностного слоя бластулы. Возникают серповидная бороздка и спинная губа бластопора.
Одновременно более мелкие клетки анимального полюса, делящиеся быстрее, начинают перемещаться в сторону вегетативного полюса. В области спинной губы они подворачиваются и впячиваются, а с боков и со стороны, противоположной серповидной бороздке, обрастают более крупные клетки. Затем образуются боковые и брюшная губы бластопора. Бластопор смыкается в кольцо, внутри которого некоторое время видны крупные светлые клетки вегетативного полюса в виде желточной пробки. Позднее они полностью погружаются внутрь, а бластопор сужается. Конкретные области бластодермы, называемые презумптивными. Известно, что у бесхвостых амфибий материал презумптивной хорды и мезодермы на стадии бластулы лежит не на ее поверхности, а во внутренних слоях стенки амфибластулы. Определенное сходство процессов гаструляции и области презумптивных органов у земноводных и ланцетника, т.е. гомология основных органов, таких, как нервная трубка, хорда, вторичная кишка, указывает на их филогенетическое родство. Гаструляция у зародышей с мepoблacтическим типом дробления и развития имеет свои особенности. К моменту откладки яйца зародыш уже состоит из нескольких слоев: верхний слой называют эпибластом, нижний-первичным гипобластом. Между ними находится узкая щель-бластоцель. Затем образуется вторичный гипобласт.
Основные события гаструляции и окончательное образование трех зародышевых листков начинаются после откладки яиц с началом инкубации. Возникает скопление клеток в задней части эпибласта. Образуется первичная полоска, которая вытягивается в направлении к головному концу. В центре первичной полоски образуется первичная бороздка, а по краям-первичные валики. На головном конце первичной полоски возникает утолщение-гензеновский узелок, а в нем -первичная ямка.
Когда клетки эпибласта входят в первичную бороздку, их форма изменяется. Они напоминают по форме «колбовидные». Затем эти клетки приобретают звездчатую форму и погружаются под эпибласт, образуя мезодерму. Энтодерма образуется на основе первичного и вторичного гипобласта с добавлением нового поколения энтодермальных клеток, мигрирующих из верхних слоев, бластодермы.
Часть клеток, мигрирующая из эпибласта через гензеновский узелок, образует будущую хорду. Одновременно с закладкой и удлинением хорды гензеновский узелок и первичная полоска постепенно исчезают в направлении от головного к хвостовому концу. Это соответствует сужению и закрытию бластопора.
Особенности стадии гаструляции. Гаструляция характеризуется разнообразными клеточными процессами. Продолжается митотическое размножение клеток. Наиболее характерная черта гаструляции состоит в перемещении клеточных масс. Это приводит к изменению строения зародыша и превращению его из бластулы в гаструлу. Происходит сортировка клеток по их принадлежности к разным зародышевым листкам, внутри которых они «узнают» друг друга.
На фазу гаструляции приходится начало цитодифференцировки, что означает переход к активному использованию биологической информации собственного генома. Одним из регуляторов генетической активности является различный химический состав цитоплазмы клеток зародыша, установившийся вследствие овоплазматической сегрегации.
Появление первичной полоски у птиц - результат индукционного взаимодействия между гипобластом и эпибластом. Изменение положения гипобласта по отношению к эпибласту вызывает изменение ориентации первичной полоски.
ЭКЗОГАСТРУЛЯЦИЯ- нарушение процесса гаструляции у зародышей многоклеточныхживотных, заключающееся в изменении обычного направления морфогенетич. движений. Происходит при неблагоприятных условиях развития; в эктодерме не дифференцируется нервная ткань, аформируется лишь атипичный эпидермис, тогда как материал хордомезодермы и энтодермы, детерминированный на предыдущих стадиях развития, претерпевает дифференцировку. Явление чётко демонстрирует необходимость контакта между хордомезодермой и эктодермой в период гаструляции у хордовых для детерминации зачатка ЦНС.
По окончании периода дробления зародыши всех многоклеточных животных вступают в период образования зародышевых слоев (листков). Этот этап принято называть гаструляцией.
В процессе гаструляции различают два этапа. Сначала образуется ранняя гаструла, имеющая два зародышевых листка: наружный – эктодерму и внутренний – энтодерму. Далее наступает поздняя гаструла, когда образуется средний зародышевый листок – мезодерма. Образование гаструлы протекает по-разному.
Выделяют 4 типа гаструляции:
1) Иммиграция — гаструляция путем выселения отдельных клеток из бластодермы внутрь. Впервые описана И. И. Мечниковым у зародышей медуз. Иммиграция должна быть униполярной, биполярной и мультиполярной, т. е. при иммиграции клетки выселяются из одной, двух или нескольких зон сразу. Иммиграция, наблюдающаяся у кишечно-полостных, стоящих в эволюционном ряду ниже всех многоклеточных, является самым древним типом гаструляции.
2) Инвагинация — гаструляция путем впячивания вегетативного полюса. Она характерна для низших хордовых, иглокожих, некоторых кишечно-полостных, ᴛ.ᴇ. она наблюдается у зародышей, развивающихся из изолецитальных яиц, характеризующихся полным равномерным дроблением.
3) Эпиболия — обрастание.
В случае если зародыш развивается из телолецитального яйца, а на вегетативном полюсе бластулы находятся крупные, богатые желтком макромеры, то прогибание вегетативного полюса затруднено, и гаструляция происходит за счёт быстрого размножения микромеров, которые обрастают вегетативный полюс. При этом макромеры оказываются внутри зародыша. Эпиболия наблюдается у земноводных, она сочетается с перемещением бластодермы внутрь зародыша (инвагинация) на границе анимального и вегетативного полюсов, т. е. эпиболия в чистом виде практически не встречается.
4) Деляминация — расслоение. При этом типе гаструляции, наблюдающейся у некоторых кишечно-полостных, имеющих бластулу в виде морулы (отсутствует в бластуле бластоцель), клетки бластодермы делятся на наружные и внутренние. В результате за счёт наружных клеток образуется эктодерма гаструлы, а за счёт внутренних — энтодерма.
ГАСТРУЛЯЦИЯ
Под гаструляцией понимают процесс превращения бластулы (двухслойного зародыша, состоящего из двух зародышевых лист-ков, наружного - эктодерма и внутреннего - энтодерма) в га- струлу. Этот процесс приводит к обособлению третьего зароды-шевого листка - мезодермы.
Гаструляция у животных всех классов представляет собой су-щественную структурную перестройку зародыша, осуществляемую путем клеточных движений и перемещений с образованием изби-рательных связей. Гаструляция обеспечивает установление основ-ного структурного плана в виде 3-зародышевых листков и даль-нейшее развитие зародыша. Поскольку гаструляция является продолжением изменений, начавшихся на стадии бластулы, то различным типам бластул соответствуют различные типы гаструл. Поэтому гаструляция у разных животных протекает различно.
Но как бы ни были разнообразны переходы бластулы в гаструлу, сущность процессов заключается в том, что происходит пе-ремещение зародышевого материала и обособление 3-зародыше-вых листков.
Перемещение клеточного материала при гаструляции осуществ-ляется путем иммиграции, инвагинации, эпиболии, деляминации.
По И. И. Мечникову наиболее древним способом гаструляции является иммиграция.
Этот способ заключается в активном высе-лении части бластомеров в бластоцель. Такая иммиграция назы-вается униполярной. Выселение бластомеров может происходить по всей поверхности бластодермы. В таком случае иммиграция называется мультиполярной. Она представляет редкое явление. Выселившиеся в бластоцель бластомеры образуют первичный внутренний зародышевый листок (энтодерму), а оставшиеся на месте бластомеры образуют первичный наружный зародышевый листок (эктодерма). Этот способ распространен у низших жи-вотных- рептилий, птиц, млекопитающих. Сама форма зародыша при иммиграции называется иммиграционной гаструлой.
Инвагинация - впячивание, наиболее простой способ гастру-ляции. Заключается во впячивании бластодермы вегетативного полюса вглубь бластоцеля. Появившееся впячивание все больше углубляется и в конце концов доходит до анимального полюса. В результате зародыш из однослойного пузырька превращается в двуслойный. Его стенка состоит из первичного наружного и пер-вичного внутреннего листков. Вследствии впячивания вегетатив-ного полюса бластоцель вытесянется. Образуется новая полость - полость первичной кишки или гастроцель. Она сообщается с внешней средой отверстием - первичным ртом или бластопором.. Края бластопора называют губами. Различают дорзальную и вент-ральную губы бластопора. Такой способ гаструляции имеет место" у ланцетника, иглокожих, низших хордовых.
Эпиболия - обрастание, способ гаструляции, при котором бы-стро делящиеся мелкие бластомеры анимального полюса (микро-меры) наползают на крупные бластомеры вегетативного полюса
(макромеры), не способные к перемещению вследствие загруженно-сти желтком. Первые образуют эктодерму, а вторые - энтодерму. Такая гаструляция характерна для амфибий.
Деляминация -расщепление, способ гаструляции, заключаю-щийся в параллельном расслоении поверхностного слоя бластоме-ров на два: первичный наружный, первичный внутренний. Такой способ гаструляции наблюдается у некоторых членистоногих и у высших позвоночных.
У некоторых^животных гаструляция может осуществляться ком-бинацией двух или более различных способов..
Читайте также:
Понятие и основные механизмы гаструляции. Типы гаструляции. Характеристика гаструляции у человека. Строение двухнедельного зародыша.
Гаструляция – процесс образования трёх зародышевых листков: экто- дерма (наружный листок), мезодерма (средний) и энтодерма (внутренний). При гаструляции происходят сложные химические и морфологические изменения, которые сопровождаются делением клеток, их ростом, перемещением и дифференцировкой.
Способы гаструляции: 1. Деламинация – расщепление на листки 2. Иммиграция – перемещение клеток вовнутрь 3. Инвагинация – впячивание пластов клеток вовнутрь 4. эпиболия – обрастание клеток Для зародыша человека характерно 2 способа гаструляции:
— деламинация — иммиграция
Первая фаза гаструляции начинается на 7- 8 сутки, во время имплантации, и осуществляется способом деламинации. При этом происходит расще-33 пление эмбриобласта на две пластинки с образованием эмбриональной энтодермы (гипобласта) и эмбриональной эктодермы (эпибласта). В состав гипобласта входит зачаток внезародышевой энтодермы. В состав эпибласта входят зачатки эктодермы, мезодермы, зародышевой энтодермы, нервной
трубки и хорды. Место соединения эпибласта и гипобласта называется зародышевым щитком. Зародышевый щиток имеет уплощённую овальную фор-
му. Вторая фаза гаструляции осуществляется на 14 – 17 сутки. Внутриматочный характер развития эмбрионов млекопитающих требует быстрого установления связи между матерью и плодом для питания плода. Именно поэтому уже на ранних стадиях, в период между 1 и 2 фазами гаструляции, появляются и быстро дифференцируются ткани, предназначенные для выполнения этой функции — провизорные органы. Провизорными, или временными, называют органы, которые развиваются в процессе эмбриогенеза вне тела зародыша и обеспечивают его рост и развитие. К провизорным органам относятся: 1) хорион 2) амнион 3) желточный мешок 4) аллантоис
Понятие дифференцировки зародышевых листков. Представление об индукции как факторе, вызывающем дифференцировку. Дифференцировка зародышевых листков и образование зачатков тканей и органов у зародыша человека.
Первым из зародышевых листков дифференцируется эктодерма. В период с 17 по 20 сутки, который называется пресомитным периодом, эктодерма подразделяется на 5 частей:
1. кожная эктодрма 2. нейроэктодерма. В пресомитном периоде здесь осуществляется про-
цесс нейруляции, т.е. вычленения из эктодермы нервной трубки и ганглиозной пластинки.
Нейруляция осуществляется последовательно через три стадии: — нервной пластинки- нервного желобка — нервной трубки и ганглиозной пластинки. Из нервной трубки в последующем развиваются головной и спинной мозг. Из ганглиозной пластинки – симпатические и парасимпатические вегетативные узлы, спинальные ганглии, мозговое вещество надпочечника, оболочки головного мозга, специальная гладкомышечная ткань
нейрального происхождения. кожная эктодерма, которая даёт начало эпидермису и производным кожи (волосы, ногти, потовые, сальные и молочные железы). 3. Прехордальная пластинка. Из неё развивается эпителий пищевода, трахеи, бронхов, лёгочных ацинусов.
4. Плакоды (слуховая, хрусталиковая) – образуют структуры, входящие
в состав органов чувств. 5. Внезародышевая эктодерма, из неё развиваются эпителий амниона и пупочного канатика Дифференцировка мезодермы Начинается на 20 сутки с головного конца зародыша, постепенно продвигаясь к каудальному концу, заканчивается на 35 сутки. Этот период называется сомитным. Каждое мезодермальное крыло дифференцируется на три части: — дорзальная часть – сомит — промежуточная часть – сегментные ножки или нефротомы — вентральная часть – спланхнотом. Дорсальная мезодерма в головном конце зародыша сегментируется на отдельные участки – сомиты. В каждом сомите различают три зоны (рис. 6):
— периферическую зону – дерматом — центральную зону – миотом — медиальную зону – склеротом. В каудальном конце зародыша дорсальная мезодерма не сегментируется и называется нефрогенной тканью. В результате образования сомитов зародыш утолщается, приподнимается и вдаётся в полость амниотического пузырька. Наступление сомитного периода сопровождается образованием туловищной складки, которая отделяет зародыш от внезародышевых органов. При формировании туловищной складки зародыш вдавливается в амнион и постепенно оказывается в нём.
Понятие и значение внезародышевых органов. Их появление в эволюции. Внезародышевые органы у человека. Образование, строение, значение амниона, желточного мешка, аллантоиса. Туловищная складка, ее образование, роль.
Развитие амниона Часть внезародышевых мезенхимных клеток выселяясь из зародышевого щитка, располагается в полости бластоцели и делит её на отдельные секторы. В результате такого расселения к гипобласту прилежит пузырёк. Эти участки составляют мезенхимные закладки других провизорных органов — амниотического пузырька и желточного мешка. Далее из эпибласта выселяются клетки, составляющие внезародышевую эктодерму, и обрастают изнутри мезенхимную закладку амниотического пузырька. В процессе роста тела эмбриона увеличивается количество жидкости в амниотическом пузырьке, и увеличиваются его размеры. На 7 неделе эмбриогенеза мезенхима наружной поверхности амниотического пузырька соединяется с внезародышевой мезенхимой хориона. С этого момента окончательно формируется амниотическая полость, ограниченная амниотической оболочкой.
Studepedia.org — это Лекции, Методички, и много других полезных для учебы материалов
Стенка амниотической оболочки состоит из двух слоёв:
1) амниотического эпителия, образовавшегося из внезародышевой эктодермы 2) соединительной ткани, образовавшейся из внезародышевой мезенхимы Та часть амниотической оболочки, которая покрывает плодную часть плаценты, называется плацентарной амниотической оболочкой, а эпителий, выстилающий эту часть – плацентарным амниотическим эпителием. Весь остальной эпителий амниотической оболочки называется внеплацентарным амниотическим эпителием. Соединительнотканная оболочка стенки амниона содержит 2 слоя: 1) плотный соединительнотканный слой, расположенный под базальной мембраной 2) рыхлый слой, состоящий из слизистой соединительной ткани, слабо связанной с соединительной тканью хориона. Таким образом, связь между амниотической оболочкой и хорионом непрочная, поэтому эти две оболочки легко отделить друг от друга. Функциональное значение плацентарного амниотического эпителия сстоит в том, что он секретирует компоненты амниотической жидкости, а вне- плацентарного – в обратном всасывании (реабсорбции) амниотической жид-36 кости
Функции амниона:
1) создание жидкой среды, в которой развивается зародыш 2) защита от механических воздействий 3) иммунная защита (в амниотической жидкости имеются Ig G и Ig A) 4) регуляторная функция. Плод постоянно заглатывает определённое количество околоплодных вод, которые стимулируют эмбриогенез и деятельность желудочно-кишечного тракта плода 5) выделительная функция. В околоплодные воды плод выделяет мочу и с ней конечные продукты обмена 6) эндокринная функция – на поздних этапах эмбриогенеза амнион вырабатывает простагландины, стимулирующие родовую деятельность.
Развитие желточного мешка Из гипобласта выселяются клетки, составляющие внезародышевую энтодерму, которые обрастают внутри мезенхимную закладку желточного мешка. На 11 день эмбриогенеза формируется желточный мешок, стенка которого состоит из внезародышевой мезенхимы и внезародышевой энтодермы.
Функции желточного мешка:
1) образование первичных кровеносных сосудов 2) кроветворная – это первый орган, в котором появляются стволовые клетки крови 3) образование стволовых половых клеток – гонобластов. Желточный мешок существует до 8 недели эмбриогенеза. После этого
он подвергается инволюции и его остатки входят в состав пупочного канатика.
Развитие аллантоиса Последним из внезародышевых органов формируется аллантоис. Его
развитию предшествует появление амниотической ножки. Амниотическая ножка – это тяж внезародышевой мезенхимы, соединяющий желточный и амниотический пузырьки с мезенхимой хориона.
Функции: 1) Участие в формировании сосудистой сети плаценты. Является проводником кровеносных сосудов из желточного мешка во вторичные ворсины. 2) Гистогенетическая – проксимальная часть аллантоиса идёт на образование части переходного эпителия мочевого пузыря. Нарушения развития аллантоиса может приводить к аномалиям этого органа.
Предыдущая29303132333435363738394041424344Следующая
По окончании периода дробления зародыши всех многоклеточных животных вступают в период образования зародышевых слоев (листков). Этот этап называется гаструляцией.
В процессе гаструляции различают два этапа. Сначала образуется ранняя гаструла, имеющая два зародышевых листка: наружный – эктодерму и внутренний – энтодерму. Затем наступает поздняя гаструла, когда образуется средний зародышевый листок – мезодерма.
Образование гаструлы протекает по-разному.
Выделяют 4 типа гаструляции:
1) Иммиграция — гаструляция путем выселения отдельных клеток из бластодермы внутрь. Впервые описана И. И. Мечниковым у зародышей медуз. Иммиграция может быть униполярной, биполярной и мультиполярной, т. е. при иммиграции клетки выселяются из одной, двух или нескольких зон сразу. Иммиграция, наблюдающаяся у кишечно-полостных, стоящих в эволюционном ряду ниже всех многоклеточных, является самым древним типом гаструляции.
2) Инвагинация — гаструляция путем впячивания вегетативного полюса. Она характерна для низших хордовых, иглокожих, некоторых кишечно-полостных, т.е. она наблюдается у зародышей, развивающихся из изолецитальных яиц, характеризующихся полным равномерным дроблением.
3) Эпиболия — обрастание.
Если зародыш развивается из телолецитального яйца, а на вегетативном полюсе бластулы находятся крупные, богатые желтком макромеры, то прогибание вегетативного полюса затруднено, и гаструляция происходит за счет быстрого размножения микромеров, которые обрастают вегетативный полюс. При этом макромеры оказываются внутри зародыша. Эпиболия наблюдается у земноводных, она сочетается с перемещением бластодермы внутрь зародыша (инвагинация) на границе анимального и вегетативного полюсов, т. е. эпиболия в чистом виде практически не встречается.
4) Деляминация — расслоение. При этом типе гаструляции, наблюдающейся у некоторых кишечно-полостных, имеющих бластулу в виде морулы (отсутствует в бластуле бластоцель), клетки бластодермы делятся на наружные и внутренние.
Занятие 4 Эмбриогенез. Дробление, гаструляция
В результате за счет наружных клеток образуется эктодерма гаструлы, а за счет внутренних — энтодерма.

Рис. 4. Типы гаструл: а – инвагинационная гаструла; б, в – две стадии развития иммиграционной гаструлы; г, д – две стадии развития деляминационной гаструлы; е, ж – две стадии развития эпиболической гаструлы; 1 – эктодерма; 2 – энтодерма; 3 – бластоцель.
Несмотря на разнообразие типов гаструляции, сущность процесса сводится к одному: однослойный зародыш (бластула) превращается в двухслойный зародыш (гаструлу).
Не нашли то, что искали? Воспользуйтесь поиском:
Читайте также:
Эмбриональный период
Дробление
Дробление - это ряд последовательных митотических делений зиготы, в результате которых огромный объем цитоплазмы яйца разделяется на многочисленные, содержащие ядра клетки меньшего размера. В результате дробления образуются клетки, которые называют бластомерами. Дробление от обычного деления отличает то, что вновь образовавшиеся бластомеры не увеличиваются в размерах. Это становится возможным вследствие выпадения пресинтетического периода интерфазы. При этом синтетический период интерфазы начинается в телофазе предшествующего митоза. Таким образом, количество бластомеров постепенно увеличивается, а их общий объем практически не изменяется. Цитоплазма клеток при дроблении делится путем возникновения впячиваний оболочки клетки (борозды дробления ).
Дробление яйцеклетки амфибий (лягушка):
1 - двуклеточная стадия; 2 - четырех-клеточная стадия; 3 - восьми-клеточная стадия;
4 - переход от восьми- к шестнад-цати-клеточной стадии (клетки анималь-ного полюса уже поделились, а клетки вегета-тивного
только начинают дробиться); 5 - более поздняя стадия дробления; 6 - бластула; 7 - бластула в разрезе.
Биологическое значение процесса дробления : благодаря повторяющимся циклам репродукции, происходит размножение генотипа зиготы; происходит накопление клеточной массы для дальнейших преобразований, зародыш из одноклеточного превращается в многоклеточный.
Деление бластомеров бывает синхронным и несинхронным . У большинства видов оно несинхронно с самого начала развития, у других становится таковым уже после первых делений.
Характер дробления определяется, прежде всего, строением яйцеклетки, главным образом, количеством желтка и особенностями его распределения в цитоплазме. В этой связи по способу дробления выделяют два основных типа яиц: полностью дробящиеся и дробящиеся частично. Полным дробление называется тогда, когда цитоплазма яйцеклетки полностью разделяется на бластомеры. Оно может быть равномерным - все образовавшиеся бластомеры имеют одинаковые размеры и форму (характерно для алецитальных и изолецитальных яйцеклеток) и неравномерным - образуются неравные по размерам бластомеры (свойственно телолецитальным яйцеклеткам с умеренным содержанием желтка). Мелкие бластомеры возникают у анимального полюса, крупные - в области вегетативного полюса зародыша.

Различные виды дробления: А - полное; Б - частичное; В - дискоидальное.
Частичное дробление - тип дробления, при котором цитоплазма яйцеклетки не полностью разделяется на бластомеры. Одним из видов частичного дробления является дискоидальное, при котором дроблению подвергается только лишенный желтка участок цитоплазмы у анимального полюса, где находится ядро. Участок цитоплазмы, подвергшийся дроблению, называется зародышевым диском . Этот тип дробления характерен для резко телолецитальных яиц с большим количеством желтка (рептилии, птицы, рыбы).
Дробление у представителей разных групп животных имеет свои особенности, однако завершается оно образованием близкой по строению структуры - бластулы.
Бластула
Бластула - однослойный зародыш. Она состоит из слоя клеток - бластодермы, ограничивающей полость - бластоцель. Бластула начинает формироваться на ранних этапах дробления благодаря расхождению бластомеров. Возникающая при этом полость заполняется жидкостью. Строение бластулы во многом зависит от типа дробления.
Целобластула (типичная бластула) образуется при равномерном дроблении. Имеет вид однослойного пузырька с большим бластоцелем (ланцетник).
Амфибластула образуется при дроблении телолецитальных яиц; бластодерма построена из бластомеров разного размера: микромеров на анимальном и макромеров на вегетативном полюсах. Бластоцель при этом смещается в сторону анимального полюса (земноводные).

Типы бластул: 1 - целобластула; 2 - амфибластула; 3 - дискобластула; 4 - бластоциста; 5 - эмбриобласт; 6 - трофобласт.
Дискобластула образуется при дискоидальном дроблении. Полость бластулы имеет вид узкой щели, находящейся под зародышевым диском (птицы).
Бластоциста представляет собой однослойный пузырек, заполненный жидкостью, в котором различают эмбриобласт (из него развивается зародыш) и трофобласт, обеспечивающий питание зародыша (млекопитающие).
Гаструла

Гаструла:
1 - эктодерма; 2 - энтодерма; 3 - бластопор; 4 - гастроцель.
После того как сформировалась бластула, начинается следующий этап эмбриогенеза - гаструляция (образование зародышевых листков). В результате гаструляции образуется двухслойный, а затем трехслойный зародыш (у большинства животных) - гаструла. Первоначально образуются наружный (эктодерма) и внутренний (энтодерма) слои. Позже между экто- и энтодермой закладывается третий зародышевый листок - мезодерма.
Зародышевые листки - отдельные пласты клеток, занимающие определенное положение в зародыше и дающие начало соответствующим органам и системам органов. Зародышевые листки возникают не только в результате перемещения клеточных масс, но и в результате дифференциации сходных между собой, сравнительно однородных клеток бластулы. В процессе гаструляции зародышевые листки занимают положение, соответствующее плану строения взрослого организма. Дифференциация - процесс появления и нарастания морфологических и функциональных различий между отдельными клетками и частями зародыша. В зависимости от типа бластулы и от особенностей перемещения клеток, различают следующие основные способы гаструляции: инвагинация, иммиграция, деламинация, эпиболия.

Типы гаструл:
1 - инвагинационная; 2 - эпиболическая; 3 - иммиграционная; 4 - деламинационная;
а - эктодерма; б - энтодерма; в - гастроцель.
При инвагинации один из участков бластодермы начинает впячиваться внутрь бластоцеля (у ланцетника). При этом бластоцель практически полностью вытесняется. Образуется двухслойный мешок, наружная стенка которого является первичной эктодермой, а внутренняя - первичной энтодермой, выстилающей полость первичной кишки, или гастроцель . Отверстие, при помощи которого полость сообщается с окружающей средой, называется бластопором , или первичным ртом . У представителей разных групп животных судьба бластопора различна. У первичноротых животных он превращается в ротовое отверстие. У вторичноротых бластопор зарастает, и на его месте нередко возникает анальное отверстие, а ротовое отверстие прорывается на противоположном полюсе (переднем конце тела).
Иммиграция - «выселение» части клеток бластодермы в полость бластоцеля (высшие позвоночные). Из этих клеток образуется энтодерма.
Деламинация встречается у животных, имеющих бластулу без бластоцеля (птицы). При таком способе гаструляции клеточные перемещения минимальны или совсем отсутствуют, так как происходит расслоение - наружные клетки бластулы преобразуются в эктодерму, а внутренние формируют энтодерму.
Эпиболия происходит, когда более мелкие бластомеры анимального полюса дробятся быстрее и обрастают более крупные бластомеры вегетативного полюса, образуя эктодерму (земноводные). Клетки вегетативного полюса дают начало внутреннему зародышевому листку - энтодерме.
Описанные способы гаструляции редко встречаются в чистом виде и обычно наблюдаются их сочетания (инвагинация с эпиболией у амфибий или деламинация с иммиграцией у иглокожих).
Чаще всего клеточный материал мезодермы входит в состав энтодермы. Он впячивается в бластоцель в виде карманообразных выростов, которые затем отшнуровываются. При образовании мезодермы происходит образование вторичной полости тела, или целома.
Процесс формирования органов в эмбриональном развитии называют органогенезом . В органогенезе можно выделить две фазы: нейруляция - образование комплекса осевых органов (нервная трубка, хорда, кишечная трубка и мезодерма сомитов), в который вовлекается почти весь зародыш, и построение остальных органов , приобретение различными участками тела типичной для них формы и черт внутренней организации, установление определенных пропорций (пространственно ограниченные процессы).
По теории зародышевых листков Карла Бэра , возникновение органов обусловлено преобразованием того или иного зародышевого листка - экто-, мезо- или энтодермы. Некоторые органы могут иметь смешанное происхождение, то есть они образованы при участии сразу нескольких зародышевых листков. Например, мускулатура пищеварительного тракта является производным мезодермы, а его внутренняя выстилка - производное энтодермы. Однако, несколько упрощая, происхождение основных органов и их систем все-таки можно связать с определенными зародышевыми листками. Зародыш на стадии нейруляции называется нейрулой . Материал, используемый на построение нервной системы у позвоночных животных, - нейроэктодерма , входит в состав дорсальной части эктодермы. Он располагается над зачатком хорды.

Нейрула:
1 - экто-дерма; 2 - хорда; 3 - вторич-ная полость тела; 4 - мезо-дерма; 5 - энто-дерма; 6 - кишеч-ная полость; 7 - нерв-ная трубка.
Сначала в области нейроэктодермы происходит уплощение клеточного пласта, что приводит к образованию нервной пластинки. Затем края нервной пластинки утолщаются и приподнимаются, образуя нервные валики.
Гаструляция у человека
В центре пластинки за счет перемещения клеток по средней линии возникает нервный желобок, разделяющий зародыш на будущие правую и левую половины. Нервная пластинка начинает складываться по средней линии. Края ее соприкасаются, а затем смыкаются. В результате этих процессов возникает нервная трубка с полостью - невроцелем .
Смыкание валиков происходит сначала в средней, а затем в задней части нервного желобка. В последнюю очередь это происходит в головной части, которая по ширине превосходит другие. Передний расширенный отдел в дальнейшем образует головной мозг, остальная часть нервной трубки - спинной. В результате нервная пластинка превращается в нервную трубку, лежащую под эктодермой.
В ходе нейруляции часть клеток нервной пластинки не входит в состав нервной трубки. Они образуют ганглиозную пластинку, или нервный гребень, - скопление клеток вдоль нервной трубки. Позднее эти клетки мигрируют по всему зародышу, образуя клетки нервных узлов, мозгового вещества надпочечников, пигментные клетки и т.п.
Из материала эктодермы, помимо нервной трубки, развиваются эпидермис и его производные (перо, волосы, ногти, когти, кожные железы и т.д.), компоненты органов зрения, слуха, обоняния, эпителий ротовой полости, эмаль зубов.
Мезодермальные и энтодермальные органы формируются не после образования нервной трубки, а одновременно с ней. Вдоль боковых стенок первичной кишки путем выпячивания энтодермы образуются карманы, или складки. Участок энтодермы, расположенный между этими складками, утолщается, прогибается, сворачивается и отшнуровывается от основной массы энтодермы. Так появляется хорда . Возникшие карманообразные выпячивания энтодермы отшнуровываются от первичной кишки и превращаются в ряд сегментарно-расположенных замкнутых мешков, называемых также целомическими мешками. Их стенки образованы мезодермой, а полость внутри представляет собой вторичную полость тела (или целом ).
Из мезодермы развиваются все виды соединительной ткани, дерма, скелет, поперечно-полосатая и гладкая мускулатура, кровеносная и лимфатическая системы, половая система.
Из энтодермы развиваются эпителий кишечника и желудка, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, секретирующие отделы передней и средней доли гипофиза, щитовидной и паращитовидной желез.

Эмбриональная индукция:
1 - зачаток хордо-мезо-дермы; 2 - полость блас-тулы; 3 - индуци-рован-ная нерв-ная трубка; 4 - инду-цирован-ная хорда; 5 - первич-ная нерв-ная трубка; 6 - пер-вич-ная хорда; 7 - форми-рование вторич-ного заро-дыша, соеди-ненного с заро-дышем-хозя-ином.
ЭМБРИОЛОГИЯ
Занятие №4
ТЕМА: «Эмбриогенез. Дробление, гаструляция»
КОНТРОЛЬНЫЕ ВОПРОСЫ
Определение понятия «дробление».
Характер дробления зиготы у человека.
Темные и светлые бластомеры, их морфологическая характеристика, потенции развития.
Морула и бластоциста, их морфологическая характеристика.
Имплантация зародыша в слизистую оболочку матки. Стадии имплантации.
Гистиотрофный и гематотрофный типы питания зародыша.
Способы гаструляции у человека.
Стадии гаструляции (деламинация и иммиграция), их сроки и механизмы.
Факторы, влияющие на механизмы гаструляции.
ЦЕЛЬ ЗАНЯТИЯ:
Усвоить особенности и характер дробления зиготы у человека. Научиться различать темные и светлые бластомеры. Усвоить различие между морулой и бластоцистой. Знать стадии имплантации и типы питания зародыша в зависимости от стадии имплантации. Изучить способы гаструляции у человека, знать стадии и механизмы гаструляции.
Студент должен знать:
характер дробления зиготы у человека;
морфологическую характеристику темных и светлых бластомеров, потенции развития;
морфологическую характеристику морулы и бластоцисты;
стадии имплантации зародыша в слизистую оболочку матки;
способы гаструляции у человека, их сроки и механизмы.
Студент должен уметь:
на схемах и таблицах уметь различать темные и светлые бластомеры; дифференцировать морулу и бластоцисту;
различать стадии имплантации зародыша в слизистую оболочку матки;
различать способы гаструляции у человека;
сделать зарисовки и составить их письменный протокол.
ДРОБЛЕНИЕ
Дробление представляет собой серию митотических делений зиготы с образованием многих дочерних клеток (бластомеров) меньшего размера. Митотические деления зиготы и в последующем бластомеров происходят с увеличением числа клеток, но без увеличения их массы, поэтому именуются дроблением.
Типы дробления:
полное, или голобластическое, (ланцетник, амфибии, млекопитающие): зигота полностью разделяется на бластомеры;
частичное, или меробластическое, (рыбы, рептилии, птицы): дробится лишь часть зиготы;
равномерное: образующиеся бластомеры одинаковы или близки по величине;
неравномерное: бластомеры отличаются по размерам;
синхронное: бластомеры делятся одновременно;
асинхронное.
По характеру пространственного расположения бластомеров выделяют следующие виды полного дробления :
радиальное: образующиеся бластомеры располагаются друг над другом, создавая фигуру с радиальной симметрией (ланцетник, амфибии);
спиральное: вышележащие бластомеры смещены по отношению к нижележащим под углом 45°, располагаясь, таким образом, по спирали (моллюски, немертины, кольчатые черви и некоторые планарии);
билатеральное (билатерально-симметричное): на ранних стадиях расположение бластомеров происходит по закону билатеральной симметрии, так что бластомерам правой стороны зародыша соответствуют точно такие же бластомеры левой стороны (асцидии, нематоды и некоторые другие беспозвоночные)
хаотическое (неупорядоченное): уже после третьего деления отсутствует строгая закономерность в расположении бластомеров, в ходе дальнейшего дробления возникающая беспорядочность усиливается (некоторые медузы). Внешняя хаотичность, однако, регулируется внутренними механизмами, которые еще не выяснены, и приводит к образованию животного конкретного вида.
Частичное дробление существует в двух видах:
дискоидальное: дробление, при котором на бластомеры делится лишь часть цитоплазмы у анимального полюса (рыбы, рептилии, птицы);
поверхностное: дробится поверхностный слой цитоплазмы, по мере развития полностью обособляющийся от желтка (насекомые и большая часть других членистоногих).
В процессе дробления зародыш перемещается по маточной трубе и заканчивает дробление в полости матки на 6-е сутки развития. Дробление завершается образованием бластулы. Это многоклеточный зародыш, как правило, с полостью внутри. Стенку бластулы называют бластодерма , а полость - бластоцель (первичная полость тела). У бластулы также обычно различают крышу, дно и разграничивающую их краевую зону . Бластулу, образующуюся при полном равномерном дроблении, называют равномерная целобластула (ланцетник), при полном неравномерном - неравномерная целобластула, называемая также амфибластулой (амфибии) или бластоцистой (млекопитающие, человек), при частичном дискоидальном - дискобластула (рептилии, птицы), при поверхностном - стерробластула (кишечнополостные).
На стадии бластулы окончательно устанавливается полярность зародыша, повышается степень его интеграции и возникает именно тот порядок расположения клеток и та степень их взаимодействия, которые необходимы для направленных клеточных движений на следующем этапе развития - гаструляции.
На стадии поздней бластулы у ланцетника и низших позвоночных устанавливаются так называемые презумптивные области, которые содержат материал определенных органов и систем. Расположение и границы презумптивных областей, или презумптивных зачатков, были изучены с помощью метода маркировки участков зародыша витальными красителями (метод В. Фогта, предложен в 1925 г.). После нанесения метки на поверхность зародыша можно проследить ее перемещения и выяснить судьбу данного участка в ходе гаструляции и дальнейшего развития.
План взаимного расположения презумптивных зачатков будущих органов получил название карты презумптивных зачатков.Карта зачатков, например, у амфибий, точно указывает положение областей, из которых будут развиваться эпидермис кожи, органы обоняния, слуха, хрусталик глаза и другие.
Дробление зародыша человека полное, или голобластическое (борозды дробления проходят через весь зародыш), неравномер ное (в результате дробления образуются бластомеры неравной величины)и асинхронное (разные бластомеры дробятся с различной скоростью, поэтому зародыш на отдельных стадиях дробления содержит нечетное число клеток). Первое дробление (деление) зиготы завершается через 30 ч, в результате образуются два бластомера, покрытых оболочкой оплодотворения, За стадией двух бластомеров следует стадия трех бластомеров.
С первых же дроблений зиготы формируются два вида бластомеров - «темные» и «светлые». «Светлые», более мелкие, бластомеры дробятся быстрее и располагаются одним слоем вокруг крупных «темных», которые оказываются в середине зародыша. Из поверхностных «светлых» бластомеров в дальнейшем возникает трофобласт, связывающий зародыш с материнским организмом и обеспечивающий его питание. Внутренние, «темные», бластомеры формируют эмбриобласт, из которого образуются тело зародыша и внезародышевые органы (амнион, желточный мешок, аллантоис).
Начиная с 3 суток, дробление идет быстрее, и на 4-е сутки зародыш состоит из 7-12 бластомеров. Через 50 - 60 ч образуется плотное скопление клеток - морула , а на 3-4-е сут. начинается формирование бластоцисты - полого пузырька, заполненного жидкостью. Бластоциста в течение 3 суток перемещается по маточной трубе к матке и через 4 суток попадает в полость матки. Бластоциста находится в полости матки в свободном виде (свободная бластоциста) в течение 2 суток (5-е и 6-е сутки). К этому времени бластоциста увеличивается в размере благодаря росту числа бластомеров - клеток эмбриобласта и трофобласта - до 100 и вследствие усиленного всасывания трофобластом секрета маточных желез и активной выработки жидкости клетками трофобласта.
Трофобласт первые 2 недели развития обеспечивает питание зародыша за счет продуктов распада материнских тканей – гистиотрофный тип питания. Эмбриобласт располагается в виде узелка зародышевых клеток («зародышевый узелок»), который прикрепляется изнутри к трофобласту на одном из полюсов бластоцисты.
ИМПЛАНТАЦИЯ
Имплантация (врастание, укоренение) - внедрение зародыша в слизистую оболочку матки. Различают две стадии имплантации:
адгезию (прилипание): зародыш прикрепляется к внутренней поверхности матки;
инвазию (погружение) - внедрение зародыша в ткани слизистой оболочки матки.
На 7-е сут. в трофобласте и эмбриобласте происходят изменения, связанные с подготовкой к имплантации:
бластоциста сохраняет оболочку оплодотворения;
в трофобласте увеличивается количество лизосом с ферментами, обеспечивающими разрушение (лизис) тканей стенки матки и тем самым способствующими внедрению зародыша в толщу ее слизистой оболочки;
появляющиеся в трофобласте микроворсинки постепенно разрушают оболочку оплодотворения;
зародышевый узелок уплощается и превращается в зародышевый щиток, в котором начинается подготовка к первой стадии гаструляции.
Имплантация продолжается около 40 часов. Одновременно с имплантацией начинается гаструляция (образование зародышевых листков). Это первый критический период развития.
В первой стадии трофобласт прикрепляется к эпителию слизистой оболочки матки, и в нем формируются два слоя - цитотрофобласт и симпластотрофобласт.
Во второй стадии симпластотрофобласт, продуцируя протеолитические ферменты, разрушает слизистую оболочку матки. Формирующиеся при этом ворсинки трофобласта, внедряясь в матку, последовательно разрушают ее эпителий, затем подлежащую соединительную ткань и стенки сосудов, и трофобласт вступает в непосредственный контакт с кровью материнских сосудов. Питание зародыша осуществляется непосредственно из материнской крови (гематотрофный тип питания). Из крови матери зародыш получает не только все питательные вещества, но и кислород, необходимый для дыхания.
Гематотрофный тип питания, сменяющий гистиотрофный, сопровождается переходом к качественно новому этапу эмбриогенеза - второй фазе гаструляции и закладке внезародышевых органов .
Гаструляция
Период гаструляции характеризуется активными перемещениями, как отдельных клеток зародыша, так и клеточных масс, в результате которых у позвоночных формируются три основных пласта тела, их называют зародышевыми листками :
эктодерма - наружный зародышевый листок;
мезодерма - средний зародышевый листок;
энтодерма - внутренний зародышевый листок.
Зародышевые листки у разных животных являются гомологичными образованиями , т.е. в ходе развития они дают идентичные структуры : эктодерма всегда преобразуется в наружный покров тела, а из энтодермы развивается выстилка средней кишки.
Другая особенность периода гаструляции заключается в том, что образующиеся при делении клетки , в отличие от бластомеров, начинают расти и увеличиваются до размеров материнской, при этом происходит активный рост и увеличение размеров самого зародыша.
Гаструляция у человека протекает в две стадии:
I стадия (деламинация) приходится на 7-е сутки внутриутробного развития: из материала зародышевого узелка (эмбриобласта) образуются два листка: наружный листок - эпибласт (клетки имеют вид псевдомногослойного призматического эпителия) и внутренний - гипобласт , обращенный в полость бластоцисты (клетки мелкие кубические, с пенистой цитоплазмой).
II стадия (иммиграция) - на 14-15-е сутки внутриутробного развития.
Способы гаструляции , т. е. механизмы образования зародышевых листков, отличаются у разных животных и определяются во многом строением бластулы. Выделяют четыре основных способа гаструляции:
инвагинация (впячивание);
деламинация (расслоение, расщепление);
иммиграция (выселение);
эпиболия (обрастание).
В результате гаструляции возникает зародыш - гаструла . Гаструла имеет полость - гастроцель (полость первичной кишки), в которую ведет отверстие - бластопор (первичный рот).
В зависимости от дальнейшей судьбы бластопора в развитии все животные подразделяются на первичноротых и вторичноротых. У первичноротых, к которым относится большинство беспозвоночных, на месте бластопора образуется ротовое отверстие, а противоположный конец становится задним концом тела. У вторичноротых, которые включают хордовых и некоторых беспозвоночных, бластопор преобразуется в анальное отверстие или нервнокишечный канал, расположенный на заднем конце тела, а ротовое отверстие прорывается на брюшной стороне у противоположного конца тела. У бластопора различают края, или губы: дорсальную, латеральные и вентральную .
Важным итогом гаструляции у хордовых является формирование в составе зародышевых листков так называемого осевого комплекса зачатков. Осевой комплекс зачатков представляет собой расположенные по оси тела зародыша зачатки нервной системы (нервная пластинка) и хорды (хордальная пластинка), а также лежащие латерально по отношению к хордальной пластинке и связанные с ней зачатки мезодермы. Тесно прилежащие друг к другу зачатки хорды и мезодермы часто называют хордомезо дермой .
Существует несколько факторов гаструляции:
физические: к таковым относится более быстрое деление мелких клеток по сравнению с более крупными, что приводит к обрастанию крупных клеток мелкими, например, в бластуле амфибий и рыб, имеющих телолецитальные яйцеклетки. В бластуле этих животных существует градиент распределения бластомеров по величине - их размеры уменьшаются по направлению от вегетативного полюса к анимальному. Такое же направление имеет метаболический градиент - с уменьшением размеров клеток повышается их метаболическая активность и темпы деления. Именно в областях активного деления клеток начинаются процессы их перемещения. Одной из причин перемещения клеток считают изменение поверхностного натяжения при увеличении числа клеток. Клетки при этом способны совершать активные амебоидные движения;
химические: механизм индуцирующего влияния связан с выделением особых факторов: белков, нуклеопротеидов, стероидов.
организующие центры: направление движения клеток, а затем и их дифференцировка определяются индукцией - влиянием одних областей или зачатков зародыша на другие. Такие области и зачатки получили название организующих центров, или индукторов. Теория организующих центров, выдвинутая Г.Шпеманом, устанавливает наличие у зародыша на разных стадиях его развития особых областей, оказывающих индуцирующее влияние на соседние участки. К организующим центрам, например, относится передняя (дорсальная) губа бластопора, индуцирующая формирование хордомезодермы, которая, в свою очередь, вызывает дифференцировку части клеток эктодермы в нервную пластинку.
Важную роль в регуляции межклеточных взаимодействий во время развития выполняют специализированные межклеточные соединения - щелевые контакты , появляющиеся уже во время дробления между бластомерами и, возможно, представляющие собой первую систему передачи сигналов между клетками.
Типы дробления (ознакомиться)
А - радиальное дробление (ланцетник);
Б - билатеральное дробление (аскарида);
В - неупорядоченное дробление (сосальщики);
Г - спиральное дробление (моллюск).
Цифрами обозначена последовательность стадий дробления.
БЕЛОРУССКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
УДК 611.06 ББК 28.03 Г 22
А в т о р ы - с о с т а в и т е л и:
Р е ц е н з е н т кандидат биологических наук,
A. М. Ходосовская
Гаструляция : метод. указания / авт.-сост.: Г. Т. Маслова,
Г 22 А. В. Сидоров. – Мн. : БГУ, 2006. – 23 с.
В данном издании изложены методические указания по курсу «Биология индивидуального развития». Предназначено для студентов биологического факультета специальности 1-31 01 01 «Биология».
УДК 611.06 ББК 28.706
Общая характеристика и определения……...………………… |
|
Способы гаструляции...………………………………………... |
|
Закладка мезодермы и формирование целома...……………... |
|
Презумптивные карты зародышей...………………………….. |
|
Зародышевые листки и их производные.……………………... |
|
Периодизация гаструляции..........………................................... |
|
Формирование осевых органов.......................................……… |
|
Причины и механизмы гаструляции………………..………… |
|
Контрольные вопросы…………………………………………. |
|
Литература……………………………………………………… |
ОБЩАЯ ХАРАКТЕРИСТИКА И ОПРЕДЕЛЕНИЯ
Гаструляция представляет собой интегрированный процесс миграции клеток, приводящий к резкому перераспределению содержимого бластулы. В результате образуются зародышевые листки:
эктодерма , энтодерма и мезодерма . Зародыш, расчлененный на зародышевые листки называется гаструлой .
Бластула состоит из большого числа клеток, локализация которых определяется еще в период дробления. Во время гаструляции эти клетки занимают новое положении и приобретают новых соседей. В этот период устанавливается план строения многослойного тела животного.
Гаструляция является промежуточным этапом единого динамического процесса, в течение которого органообразующие участки бластулы перестраиваются так, чтобы затем было легче перейти к органогенезу и сформировать организм.
Клетки, которые в будущем образуют энтодермальные и мезодермальные органы попадают внутрь зародыша, тогда как клетки из которых возникнут кожа и нервная система распространяются по его поверхности (органы эктодермального происхождения). Таким образом, три зародышевых листка – наружная эктодерма, внутренняя энтодерма и лежащая между ними мезодерма впервые формируются в процессе гаструляции. При этом создаются условия для взаимодействия между ними.
Гаструляция, с которой связаны возникновение различий в клетках и самая ранняя дифференцировка зародыша, сопряжена с экспрессией генов зародыша. Дифференциальную активность генов в процессе гаструляции отражают понятия компетенции и детерминации.
Компетентность – это способность клетки дифференцироваться в нескольких, немногих, направлениях.
Детерминация – это состояние, при котором клетка уже вступила на путь определенной дифференциации и находится в самом его начале.
Согласно ныне существующему определению (Ч. Бодемер, 1971), гаструляция представляет собой ряд морфогенетических движений, в результате которых проспективные зачатки тканей (эктодерма, мезодерма и энтодерма) перемещаются в места, предназначенные для них в соответствии с планом организации.
СПОСОБЫ ГАСТРУЛЯЦИИ
Характер гаструляции в животном царстве необычайно сильно варьирует, однако осуществляется она при участии сравнительно

немногочисленных механизмов. При гаструляции наблюдается сочетание нескольких основных типов движений.
инвагинация
Представляет собой вворачивание участка клеточной стенки зародыша подобно тому, как вдавливается внутрь мягкая стенка резинового мяча, когда на него нажимают. При инвагинации механическая целостность стенки бластулы не нарушается. Впячивание одного из участков бластодермы внутрь осуществляется целым пластом
эктодерма
энтодерма
гастроцель
Рис. 1. Схема процесса инвагинации:
А – движение пласта клеток; Б – инвагинационная гаструла
У ланцетника впячиваются клетки вегетативного полюса, а у земноводных инвагинация происходит на границе между анимальным и вегетативным полюсом в области серого серпа . Процесс инвагинации возможен только в яйцах с небольшим или средним количеством желтка.
В результате образуется двуслойный мешок, наружной стенкой которого является первичная эктодерма, а внутренней – первичная энтодерма. Бластоцель (первичная полость тела или полость Бэра) постепенно вытесняется, в ходе впячивания образуется гастроцель - первичный кишечник или архентерон , а отверстие при помощи которого он сообщается с внешней средой, называют бластопором или первичным ртом . Последний со всех сторон окружен губами бластопора .
Судьба бластопора у разных животных неодинакова. У многих животных первичный рот, развиваясь и дифференцируясь, превращается
в дефинитивный рот взрослого организма - первичноротые (черви, моллюски и членистоногие). Не менее обширна группа вторичноротых , у которых бластопор превращается в анальное отверстие (щетинкочелюстные, плеченогие, иглокожие, кишечнодышащие) или в нервно-кишечный канал, находящийся в заднем конце эмбриона (хордовые).

инволюция
При этом происходит вворачивание внутрь зародыша увеличивающегося в размерах наружного пласта клеток, который распространяется по внутренней поверхности остающихся снаружи клеток (рис. 2).
эктодерма |
|
энтодерма |
|
Рис. 2. Схема инволюции:
А - исходный пласт клеток, Б – движения клеточного пласта в ходе инволюции
иммиграция
Представляет собой наиболее древний способ гаструляции, открытый И. И. Мечниковым (1886). Иммиграционная гаструла характерна для многих кишечнополостных. Она развивается вследствие активного выселения (ингрессии ) части клеток стенки бластулы внутрь бластоцеля. Это выселение клеток может происходить как с одного (вегетативного) полюса – униполярная иммиграция, так и с двух противоположных (анимального и вегетативного) полюсов – биполярная иммиграция (рис. 3).
эктодерма
Выселяющиеся в полость бластулы клетки вегетативного полюса (образуют энтодерму)
Рис. 3. Униполярная иммиграция (схема)
Иногда процессы иммиграции идут без особого порядка по всей поверхности бластулы - мультиполярная иммиграция. Выселившиеся клетки позднее образуют внутренний слой гаструлы – энтодерму. Таким образом, как и в ходе инвагинации зародыш становится двухслойным.
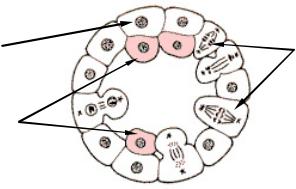
Униполярная иммиграция характерна почти для всех гидромедуз, особенно тех, которым свойственна плавающая целобластула. Би- и мультиполярная иммиграция, напротив, встречаются гораздо реже. У многих кишечнополостных, которым свойственна иммиграционная гаструла, происходит столь массовое, активное выселение клеток бластодермы, что бластоцель полностью заполняется их плотной массой и исчезает. Важной особенностью иммиграционной гаструлы является отсутствие бластопора, а значит нет и характерного для инвагинационной гаструлы сообщения гастроцеля с внешней средой.
При иммиграции происходит перемещение отдельных клеток или групп клеток, не объединенных в единый пласт. Иммиграция, в том или ином виде, встречается у всех зародышей, но в наибольшей степени характерна для второй фазы гаструляции высших позвононых (птиц и млекопитающих).
деляминация
В тех случаях, когда дробление заканчивается образованием бластулы с невыраженной или почти отсутствующей полостью (бластоцелем), например морулы у некоторых кишечнополостных, наблюдается расщепление единого клеточного пласта на два более или менее параллельных. Каждая клетка, образующая бластодерму, путем
Рис. 4. Деляминация (схема)
При деляминации клеточные перемещения практически отсутствуют. Этот тип гаструляции ограничивается выравниванием внутренних стенок клеток наружного слоя, причем такое выравнивание нередко идет волной от одной соседней клетки к другой. Вдоль выровненных поверхностей формируется базальная мембрана, отделяющая внешний клеточный слой (эктодерму) от внутренней массы

клеток, которая вся становится энтодермой. Деляминацию можно наблюдать у зародышей с неполным типом дробления, таких как пресмыкающиеся, птицы, низшие млекопитающие.
эпиболия
Представляет движение эпителиальных пластов клеток (обычно эктодермальных), которые распространяются как одно целое, а не индивидуально и окружают глубокие слои зародыша. Другими словами, происходит обрастание мелкими клетками анимального полюса более крупных, отстающих в скорости деления и менее подвижных, клеток вегетативного полюса (рис. 5).
Рис. 5. Эпиболия (схема)
Формирование эпиболической гаструлы ярко выражено у земноводных и некоторых организмов с резко телолецитальным строением яиц. В результате у зародышей таких животных бластопор отсутствует и архентерон не формируется. Только впоследствие, когда макромеры делясь становятся меньшего размера, образуется полость - формируется зачаток первичного кишечника.
В чистом виде указанные способы гаструляции встречаются крайне редко. В каждом конкретном случае эмбриогенеза, как правило, сочетаются несколько типов движений. В результате принято выделять последний, смешанный , способ гаструляции.
ЗАКЛАДКА МЕЗОДЕРМЫ И ФОРМИРОВАНИЕ ЦЕЛОМА
После завершения гаструляции материал, оставшийся на поверхности зародыша, формирует наружный зародышевый листок – эктодерму, а погрузившийся любым способом внутренний зародышевый листок (энтодерма) образует впоследствии стенку пищеварительного тракта с его производными. Кроме того, погрузившийся внутрь материал
помимо энтодермы содержит и закладку среднего зародышевого листка
– мезодермы , который потом отделяется от энтодермы.
Таким образом, в связи с гаструляцией, параллельно с ней или на следующем, обусловленном ей же, этапе у всех животных, за исключением кишечнополостных, возникает мезодерма. Это совокупность клеточных элементов залегающих между эктодермой и энтодермой, т.е. в бластоцеле. В результате зародыш становится не двуслойным, а трехслойным. У высших позвоночных трехслойное строение зародышей возникает уже в процессе гаструляции, тогда как у низших хордовых и у всех других типов в результате собственно гаструляции образуется лишь двуслойный эмбрион.
В настоящее время вопросы образования мезодермы у животных, в общем, могут считаться решенными в плоскости формальноморфологического описания этих процессов. Если отвлечься от разнообразных деталей формирования мезодермы у разных животных, то можно выделить два принципиально разных пути ее образования.
телобластический
У первичноротых во время гаструляции на границе между эктодермой и энтодермой, по бокам бластопора, уже имеются две большие клетки (или несколько таких клеток) – телобласты, получившие в ходе делений дробления всю полярную ооплазму. Впоследствие они отделяют от себя мелкие клетки и формируют, таким образом, средний пласт – мезодерму. Постепенно телобласты, давая новые и новые поколения клеток мезодермы, отодвигаются к заднему концу зародыша. По этой причине такой способ закладки называют телобластическим.
Телобластический способ закладки мезодермы встречается, преимущественно, у спирально дробящихся форм. В результате, из потомков 2d и 4d бластомеров возникает пара так называемых мезодермальных полосок . Позже они подразделяются на парные структуры - сомиты , внутри которых путем расхождениия клеток образуются участки вторичной полости тела или целома . Такой способ образования полостей называется шизоцельным или кавитационным . В результате такого способа закладки, мезодерма никак не связана с энтодермой, образующейся из других бластомеров. Закладка мезодермы из отдельных, предназначенных к тому бластомеров, имеет также место у большинства круглых червей, некоторых ракообразных и в ряде других групп первичноротых животных.
энтероцельный
Принципиально другой способ закладки мезодермы наблюдается у вторичноротых животных: иглокожих, бесчерепных (ланцетника),
кишечнодышаших, плеченогих, хордовых. Материал будущей мезодермы вворачивается вместе с энтодермой в составе единого гастрального впячивания, и в процессе инвагинации граница между обеими закладками, как правило, не различима. Прослеживая судьбу закладок в обратном, ретроспективном порядке, от поздних стадий развития назад к ранним, можно выяснить какую часть гастрального впячивания выстилает материал будущей мезодермы. Такое впячивание, включающее в себя материал и энтодермы, и мезодермы, а у хордовых еще и хорды, называется первичным кишечником, тогда как гастроцель представляет собой полость первичной кишки.
Мезодерма выселяется из архентерона:
- путем выпячивания его стенок и отшнуровки возникших выпячиваний;
- путем деламинации стенки архентерона;
- путем иммиграции клеток из стенки архентерона.
После отделения мезодермы в составе стенки архентерона остается уже чисто энтодермальный материал и архентерон превращается в полость дефинитивной кишки. Полость отшнуровавшихся мезодермальных карманов – это целом.
У всех животных, которым свойственна вторичная полость тела, начало целомическим мешкам дает мезодерма. При энтероцельном происхождении мезодермы целомические мешки образуются путем изменения и последующей дифференцировки карманоподобных выпячиваний архентерона, а при телобластическом способе образования
– за счет расхождения мезодермальных тяжей. В обоих случаях целомические мешки формируются симметрично по бокам кишечника.
Таким образом, в ходе развития зародыша формируются различные полости, имеющие важное морфогенетическое или функциональное значение. Сначала появляется полость бластулы. Затем в связи с процессами гаструляции возникает гастроцель, наконец у многих животных вторичная полость тела – целом. При образовании гастроцели и целома бластоцель все больше уменьшается, так что от нее остаются лишь щели в промежутках между стенками кишки и целома. Эти щели у некоторых животных превращаются в полости кровеносной системы. Гастроцель же со временем превращается в полость средней кишки.
Процессы гаструляции непосредственно предшествуют периоду органогенеза. У одних животных закладываются органы и системы органов, приобретающие постепенно дефинитивное значение, у других животных формируются сначала органы, свойственные личинке, а затем совершается метаморфоз и происходят процессы формирования
Сущность стадии гаструляции заключается в том, что однослойный зародыш - бластула - превращается в многослойный - двух- или трехслойный, называемый гаструлой (от греч. гастер - желудок в уменьшительном смысле).
У примитивных хордовых, например у ланцетника, однородная однослойная бластодерма во время гаструляции преобразуется в наружный зародышевый листок -эктодерму -и внутренний зародышевый листок - энтодерму. Энтодерма формирует первичную кишку с полостью внутри-гастроцель. Отверстие, ведущее в гастроцель, называют бластопором или первичным ртом. Два зародышевых листка являются определяющими морфологическими признаками гаструляции. Их существование на определенной стадии развития у всех многоклеточных животных, начиная с кишечнополостных и кончая высшими позвоночными, позволяет думать о гомологии зародышевых листков и единстве происхождения всех этих животных.
У позвоночных помимо двух упомянутых во время гаструляции образуется еще третий зародышевый листок - мезодерма, занимающая место между экто- и энтодермой. Развитие среднего зародышевого листка, представляющего собой хордомезодерму, является эволюционным усложнением фазы гаструляции у позвоночных и связано с ускорением у них развития на ранних стадиях эмбриогенеза. У более примитивных хордовых животных, таких, как ланцетник, хордомезодерма обычно образуется в начале следующей после гаструляции фазы - органогенезе. Смещение времени развития одних органов относительно других у потомков по сравнению с предковыми группами является проявлением гетерохронии. Изменение времени закладки важнейших органов в процессе эволюции встречается не редко.
Процесс гаструляции характеризуется важными, клеточными преобразованиями, такими, как направленные перемещения групп и отдельных клеток, избирательное размножение и сортировка клеток, начало цитодифференцировки и индукционных взаимодействий.
Способы гаструляции различны . Выделяют четыре разновидности направленных в пространстве перемещений клеток, приводящих к преобразованию зародыша из однослойного в многослойный.
Инвагинация - впячивание одного из участков бластодермы внутрь целым пластом. У ланцетника впячиваются клетки вегетативного полюса, у земноводных инвагинация происходит на границе между анимальным и вегетативным полюсами в области серого серпа. Процесс инвагинации возможен только в яйцах с небольшим или средним количеством желтка.
Эпиболия - обрастание мелкими клетками анимального полюса более крупных, отстающих в скорости деления и менее подвижных клеток вегетативного полюса. Такой процесс ярко выражен у земноводных.
Деламинация -расслоение клеток бластодермы на два слоя, лежащих друг над другом. Деламинацию можно наблюдать в дискобластуле зародышей с частичным типом дробления, таких, как пресмыкающиеся, птицы, яйцекладущие млекопитающие. Деламинация проявляется в эмбриобласте плацентарных млекопитающих, приводя к образованию гипобласта и эпибласта.
Иммиграция - перемещение групп или отдельных клеток, не объединенных в единый пласт. Иммиграция встречается у всех зародышей, но в наибольшей степени характерна для второй фазы гаструляции высших позвоночных.
В каждом конкретном случае эмбриогенеза, как правило, сочетаются несколько способов гаструляции.
Особенности стадии гаструляции.
Гаструляция характеризуется разнообразными клеточными процессами. Продолжается митотическое размножение клеток, причем оно имеет разную интенсивность в разных частях зародыша. Вместе с тем наиболее характерная черта гаструляции состоит в перемещении клеточных масс. Это приводит к изменению строения зародыша и превращению его из бластулы в гаструлу. Происходит сортировка клеток по их принадлежности к разным зародышевым листкам, внутри которых они «узнают» друг друга.На фазу гаструляции приходится начало цитодифференцировки, что означает переход к активному использованию биологической информации собственного генома. Одним из регуляторов генетической активности является различный химический состав цитоплазмы клеток зародыша, установившийся вследствие овоплазматической сегрегации. Так, эктодермальные клетки земноводных имеют темный цвет из-за пигмента, попавшего в них из анимального полюса яйцеклетки, а клетки энтодермы - светлый, так как происходят из вегетативного полюса яйца.Во время гаструляции очень велика роль эмбриональной индукции. Показано, что появление первичной полоски у птиц - результат индукционного взаимодействия между гипобластом и эпибластом. Гипобласту присуща полярность. Изменение положения гипобласта по отношению к эпибласту вызывает изменение ориентации первичной полоски.Такие проявления целостности зародыша как детерминация, эмбриональная регуляция и интегрированность присущи ему во время гаструляции в той же мере как и во время дробления
30. Первичный органогенез (нейруляция) как процесс образования комплекса осевых органов хордовых. Дифференцировка зародышевых листков. Образование органов и тканей.
Органогенезы, заключающиеся в образовании отдельных органов, составляют основное содержание эмбрионального периода. Они продолжаются в личиночном и завершаются в ювенильном периоде. Органогенезы отличаются наиболее сложными и разнообразными морфогенетическими преобразованиями. Необходимой предпосылкой перехода к органогенезам является достижение зародышем стадии гаструлы, а именно формирование зародышевых листков. Занимая определенное положение друг по отношению к Другу, зародышевые листки, контактируя и взаимодействуя, обеспечивают такие взаимоотношения между различными клеточными группами, которые стимулируют их развитие в определенном направлении. Это так называемая эмбриональная индукция -важнейшее следствие взаимодействия между зародышевыми листками.
В ходе органогенезов изменяются форма, структура и химический состав клеток, обособляются клеточные группы, представляющие собой зачатки будущих органов. Постепенно развивается определенная форма органов, устанавливаются пространственные и функциональные связи между ними. Процессы морфогенеза сопровождаются дифференциацией тканей и клеток, а также избирательным и неравномерным ростом отдельных органов и частей организма. Обязательным условием органогенезов наряду с размножением, миграцией и сортировкой клеток является их избирательная гибель.
Самое начало органогенеза называют нейруляцией . Нейруляция охватывает процессы от появления первых признаков формирования нервной пластинки до замыкания ее в нервную трубку.Параллельно формируются хорда и вторичная кишка, а лежащая по бокам от хорды мезодерма расщепляется в краниокаудальном направлении на сегментированные парные структуры - сомиты.
Нервная система позвоночных, включая человека, отличается устойчивостью основного плана строения на протяжении всей эволюционной истории подтипа. В формировании нервной трубки у всех хордовых много общего. Вначале неспециализированная спинная эктодерма , отвечая на индукционное воздействие со стороны хордомезодермы, превращается в нервную пластинку , представленную нейроэпителиальными клетками цилиндрической формы.
Нервная пластинка недолго остается уплощенной. Вскоре ее боковые края приподнимаются, образуя нервные валики, которые лежат по обе стороны неглубокой продольной нервной бороздки. Края нервных валиков далее смыкаются, образуя замкнутую нервную трубку с каналом внутри - невроцелем. Раньше всего смыкание нервных валиков происходит на уровне начала спинного мозга, а затем распространяется в головном и хвостовом направлениях. Показано, что в морфогенезе нервной трубки большую роль играют микротрубочки и микрофиламенты нейроэпителиальных клеток. Разрушение этих клеточных структур колхицином и цитохалазином В приводит к тому, что нервная пластинка остается открытой. Несмыкание нервных валиков ведет к врожденным порокам развития нервной трубки.
После смыкания нервных валиков клетки, первоначально располагавшиеся между нервной пластинкой и будущей кожной эктодермой, образуют нервный гребень . Клетки нервного гребня отличаются способностью к обширным, но строго регулируемым миграциям по всему телу и образуют два главных потока. Клетки одного из них-поверхностного -включаются в эпидермис или дерму кожи, где дифференцируются в пигментные клетки. Другой поток мигрирует в брюшном направлении , образует чувствительные спинномозговые ганглии, симпатические нервные узлы, мозговое вещество надпочечников, парасимпатические ганглии. Клетки из черепного отдела нервного гребня дают начало как нервным клеткам, так и ряду других структур, таких, как жаберные хрящи, некоторые кроющие кости черепа.
Мезодерма , занимающая место по бокам от хорды и распространяющаяся далее между кожной эктодермой и энтодермой вторичной кишки, подразделяется на дорсальную и вентральную области. Дорсальная часть сегментирована и представлена парными сомитами. Закладка сомитов идет от головного к хвостовому концу. Вентральная часть мезодермы, имеющая вид тонкого слоя клеток, называется боковой пластинкой . Сомиты соединены с боковой пластинкой промежуточной мезодермой в виде сегментированных ножек сомитов.
Все области мезодермы постепенно дифференцируются. В начале формирования сомиты имеют конфигурацию, характерную для эпителия с полостью внутри. Под индукционным воздействием, исходящим от хорды и нервной трубки, вентромедиальные части сомитов - склеротомы -превращаются во вторичную мезенхиму, выселяются из сомита и окружают хорду и вентральную часть нервной трубки. В конце концов из них образуются позвонки, ребра и лопатки.
Дорсолатеральная часть сомитов с внутренней стороны образуетмиотомы , из которых разовьются поперечно-полосатые скелетные мышцы тела и конечностей. Наружная дорсолатеральная часть сомитов образует дерматомы , которые дают начало внутреннему слою кожи - дерме. Из области ножек сомитов с зачатками нефротом и гонотом образуются органы выделения и половые железы.
Правая и левая несегментированные боковые пластинки расщепляются на два листка, ограничивающих вторичную полость тела - целом . Внутренний листок , прилежащий к энтодерме, называют висцеральным. Он окружает кишку со всех сторон и образует брыжейку, покрывает легочную паренхиму и мышцу сердца. Наружный листок боковой пластинки прилежит к эктодерме и называется париетальным. В дальнейшем он образует наружные листки брюшины, плевры и перикарда.
Энтодерма у всех зародышей в конечном счете образует эпителий вторичной кишки и многие ее производные. Сама вторичная кишка всегда располагается под хордой.
Таким образом, в процессе нейруляции возникает комплекс осевых органов нервная трубка - хорда - кишка, представляющих собой характернейшую черту организации тела всех хордовых. Одинаковое происхождение, развитие и взаимное расположение осевых органов выявляют их полную гомологию и эволюционную преемственность.
При углубленном рассмотрении и сравнении процессов нейруляции у конкретных представителей типа хордовых выявляются некоторые различия, которые связаны в основном с особенностями, зависящими от строения яйцеклеток, способа дробления и гаструляции.Обращают внимание отличающаяся форма зародышей и смещение времени закладки осевых органов друг относительно друга, т.е. описанная выше гетерохрония.
Эктодерма, мезодерма и энтодерма в ходе дальнейшего развития, взаимодействуя друг с другом, участвуют в формировании определенных органов. Возникновение зачатка органа связано с местными изменениями определенного участка соответствующего зародышевого листка. Так, из эктодермы развиваются эпидермис кожи и его производные (перо, волосы, ногти, кожные и молочные железы), компоненты органов зрения; слуха, обоняния, эпителий ротовой полости, эмаль зубов. Важнейшими эктодермальными производными являются нервная трубка, нервный гребень и образующиеся из них все нервные клетки.
Производными энтодермы являются эпителий желудка и кишки, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, а также секретирующие клетки передней и средней долей гипофиза, щитовидной и паращитовидной желез.
Мезодерма помимо уже описанных выше скелетных структур, скелетной мускулатуры, дермы кожи, органов выделительной и половой систем образует сердечно-сосудистую систему, лимфатическую систему, плевру, брюшину и перикард. Из мезенхимы, имеющей смешанное происхождение за счет клеток трех зародышевых листков, развиваются все виды соединительной ткани, гладкая мускулатура, кровь и лимфа.
Зачаток конкретного органа формируется первоначально из определенного зародышевого листка, но затем орган усложняется и в итоге в его формировании принимают участие два или три зародышевых листка.
31. Провизорные органы хордовых. Группа Анамнии и Амниоты. Образование, строение, особенности функционирования и эволюции провизорных органов и зародышевых оболочек. Амнион, хорион или сероза, аллантоис, желточный мешок, плацента. Типы плаценты, ее значение.
У животных разных типов в период эмбрионального развития возникают провизорные эмбриональные органы , обеспечивающие жизненно важные функции: дыхание, питание, выделение, движение и др. Недоразвитые органы самого зародыша еще не способны функционировать по назначению, хотя обязательно играют какую-то роль в системе развивающегося целостного организма. Как только зародыш достигает необходимой степени зрелости, когда большинство органов способны выполнять жизненно важные функции, временные органы рассасываются или отбрасываются.
Время образования провизорных органов зависит от того, какие запасы питательных веществ были накоплены в яйцеклетке и в каких условиях среды происходит развитие зародыша. У бесхвостых земноводных, например, благодаря достаточному количеству желтка в яйцеклетке и тому, что развитие идет в воде, зародыш осуществляет газообмен и выделяет продукты диссимиляции непосредственно через оболочки яйца и достигает стадии головастика. На этой стадии образуются провизорные органы дыхания (жабры), пищеварения и движения, приспособленные к водному образу жизни. Перечисленные личиночные органы дают возможность головастику продолжить развитие. По достижении состояния морфофункциональной зрелости органов взрослого типа временные органы исчезают в процессе метаморфоза.
У пресмыкающихся и птиц запасов желтка в яйцеклетке больше, но развитие идет не в воде, а на суше. В связи с этим очень рано возникает потребность в обеспечении дыхания и выделения, а также в защите от высыхания. У них уже в раннем эмбриогенезе, почти параллельно с нейруляцией, начинается формирование провизорных органов, таких, как амнион, хорион и желточный мешок. Чуть позднее формируется аллантоис. У плацентарных млекопитающих эти же провизорные органы образуются еще раньше, поскольку в яйцеклетке очень мало желтка. Развитие таких животных происходит внутриутробно, образование провизорных органов у них совпадает по времени с периодом гаструляции.
Наличие или отсутствие амниона и других провизорных органов лежит в основе деления позвоночных на две группы: Amniota и Anamnia. Эволюционно более древние позвоночные, развивающиеся исключительно в водной среде и представленные такими классами, как Круглоротые, Рыбы и Земноводные, не нуждаются в дополнительных водных и других оболочках зародыша и составляют группу анамний. К группе амниот относят первичноназемных позвоночных, т.е. тех, у кого эмбриональное развитие протекает в наземных условиях.
Это три класса: Пресмыкающиеся, Птицы и Млекопитающие. Они являются высшими позвоночными, так как имеют скоординированные и высокоэффективные системы органов, обеспечивающие им существование в наиболее сложных условиях, каковыми являются условия суши. Эти классы насчитывают большое количество видов, вторично перешедших в водную среду. Таким образом, высшие позвоночные оказались в состоянии освоить все среды обитания. Подобное совершенство было бы невозможным, в том числе и.без внутреннего осеменения и специальных провизорных эмбриональных органов.
В строении и функциях провизорных органов различных амниот много общего. Характеризуя в самом общем виде провизорные органы зародышей высших позвоночных, называемые также зародышевыми оболочками, следует отметить, что все они развиваются из клеточного материала уже сформировавшихся зародышевых листков. Некоторые особенности имеются в развитии зародышевых оболочек плацентарных млекопитающих, о чем будет сказано ниже.
· Амнион представляет собой эктодермальный мешок, заключающий зародыша и заполненный амниотической жидкостью. Амниотическая оболочка специализирована для секреции и поглощения амниотической жидкости, омывающей зародыш. Амнион играет первостепенную роль в защите зародыша от высыхания и от механических повреждений, создавая для него наиболее благоприятную и естественную водную среду. Амнион имеет и мезодермальный слой из внезародышевой соматоплевры, который дает начало гладким мышечным волокнам. Сокращения этих мышц вызывают пульсацию амниона, а медленные колебательные движения, сообщаемые при этом зародышу, по-видимому, способствуют тому, что его растущие части не мешают друг другу.
· Хорион (сероза) - самая наружная зародышевая оболочка, прилежащая к скорлупе или материнским тканям, возникающая, как и амнион, из эктодермы и соматоплевры. Хорион служит для обмена между зародышем и окружающей средой. У яйцекладущих видов основная его функция - дыхательный газообмен; у млекопитающих он выполняет гораздо более обширные функции, участвуя помимо дыхания в питании, выделении, фильтрации и синтезе веществ, например гормонов.
· Желточный мешок имеет энтодермальное происхождение, покрыт висцеральной мездермой и непосредственно связан с кишечной трубкой зародыша. У зародышей с большим количеством желтка он принимает участие в питании. У птиц, например в спланхноплевре желточного мешка, развивается сосудистая сеть. Желток не проходит через желточный проток, соединяющий мешок с кишкой. Сначала он переводится в растворимую форму под действием пищеварительных ферментов, продуцируемых энтодермальными клетками стенки мешка. Затем попадает в сосуды и с кровью разносится по всему телу зародыша.У млекопитающих нет запасов желтка и сохранение желточного мешка может быть связано с важными вторичными функциями. Энтодерма желточного мешка служит местом образования первичных половых клеток, мезодерма дает форменные элементы крови зародыша. Кроме того, желточный мешок млекопитающих заполнен жидкостью, отличающейся высокой концентрацией аминокислот и глюкозы, что указывает на возможность обмена белков в желточном мешке. Судьба желточного мешка у разных животных несколько различна. У птиц к концу периода инкубации остатки желточного мешка уже находятся внутри зародыша, после чего он быстро исчезает и к концу 6-х суток после вылупления полностью рассасывается. У млекопитающих желточный мешок бывает развит по-разному. У хищников он сравнительно большой, с сильно развитой сетью сосудов, а у приматов быстро сморщивается и исчезает без остатка до родов.
· Аллантоис развивается несколько позднее других внезародышевых органов. Он представляет собой мешковидный вырост вентральной стенки задней кишки. Следовательно, он образован энтодермой изнутри и спланхноплеврой снаружи. У рептилий и птиц аллантоис быстро дорастает до хориона и выполняет несколько функций. Прежде всего это вместилище для мочевины и мочевой кислоты, которые представляют собой конечные продукты обмена азотсодержащих органических веществ. В аллантоисе хорошо развита сосудистая сеть, благодаря чему вместе с хорионом он участвует в газообмене. При вылуплении наружная часть аллантоиса отбрасывается, а внутренняя - сохраняется в виде мочевого пузыря.
У многих млекопитающих аллантоис тоже хорошо развит и вместе с хорионом образует хориоаллантоисную плаценту. Термин плацента означает тесное наложение или слияние зародышевых оболочек с тканями родительского организма. У приматов и некоторых других млекопитающих энтодермальная часть аллантоиса рудиментарна, а мезодермальные клетки образуют плотный тяж, протягивающийся от клоакального отдела к хориону. По мезодерме аллантоиса к хориону растут сосуды, посредством которых плацента выполняет выделительную, дыхательную и питательную функции.
Плаценты различаются по форме и размещению ворсинок. По этому признаку выделяют следующие типы плацент . Диффузная – вся поверхность плодного пузыря равномерно покрыта ворсинками. Такая плацента характерна для свиньи. У жвачных животных наблюдается котиледонная плацента, где ворсинки собраны в группы – котиледоны. Поясная плацента характерна для хищных млекопитающих. В этом случае ворсинки окружают плодный пузырь в форме широкого пояса. Следующий тип плаценты – дискоидальная. Она наблюдается у обезьян и человека, когда ворсинки располагаются на плодном пузыре в виде диска.
Плацента имеет громадное значение для развивающегося детеныша.
Она выполняет ряд важных функций:
1) трофическую – посредством плаценты происходит питание плода;
2) дыхательную – осуществляет поступление кислорода;
3) экскреторную – происходит выделение в кровь материнского организма продуктов обмена;
4) защитную – предохраняет зародыш от проникновения различных бактерий;


