Жаропонижающие средства для детей назначаются педиатром. Но бывают ситуации неотложной помощи при лихорадке, когда ребенку нужно дать лекарство немедленно. Тогда родители берут на себя ответственность и применяют жаропонижающие препараты. Что разрешено давать детям грудного возраста? Чем можно сбить температуру у детей постарше? Какие лекарства самые безопасные?
ГЕНЕТИЧЕСКИЙ КОД, система записи наследственной информации в виде последовательности оснований нуклеотидов в молекулах ДНК (у некоторых вирусов - РНК), определяющая первичную структуру (расположение аминокислотных остатков) в молекулах белков (полипептидов). Проблема генетического кода была сформулирована после доказательства генетической роли ДНК (американские микробиологи О. Эйвери, К. Мак-Леод, М. Маккарти, 1944) и расшифровки её структуры (Дж. Уотсон, Ф. Крик, 1953), после установления того, что гены определяют структуру и функции ферментов (принцип «один ген - один фермент» Дж. Бидла и Э. Тейтема, 1941) и что существует зависимость пространственной структуры и активности белка от его первичной структуры (Ф. Сенгер, 1955). Вопрос о том, как комбинации из 4 оснований нуклеиновых кислот определяют чередование 20 обычных аминокислотных остатков в полипептидах, впервые поставил Г. Гамов в 1954 году.
На основании эксперимента, в котором исследовали взаимодействия вставок и выпадений пары нуклеотидов, в одном из генов бактериофага Т4 Ф. Крик и другие учёные в 1961 году определили общие свойства генетического кода: триплетность, т. е. каждому аминокислотному остатку в полипептидной цепи соответствует набор из трёх оснований (триплет, или кодон) в ДНК гена; считывание кодонов в пределах гена идёт с фиксированной точки, в одном направлении и «без запятых», то есть кодоны не отделены какими-либо знаками друг от друга; вырожденность, или избыточность, - один и тот же аминокислотный остаток могут кодировать несколько кодонов (кодоны-синонимы). Авторы предположили, что кодоны не перекрываются (каждое основание принадлежит только одному кодону). Прямое изучение кодирующей способности триплетов было продолжено с использованием бесклеточной системы синтеза белка под контролем синтетической матричной РНК (мРНК). К 1965 году генетический код был полностью расшифрован в работах С. Очоа, М. Ниренберга и Х. Г. Кораны. Раскрытие тайны генетического кода явилось одним из выдающихся достижений биологии в 20 веке.
Реализация генетического кода в клетке происходит в ходе двух матричных процессов - транскрипции и трансляции. Посредником между геном и белком является мРНК, образующаяся в процессе транскрипции на одной из нитей ДНК. При этом последовательность оснований ДНК, несущая информацию о первичной структуре белка, «переписывается» в виде последовательности оснований мРНК. Затем в ходе трансляции на рибосомах последовательность нуклеотидов мРНК считывается транспортными РНК (тРНК). Последние имеют акцепторный конец, к которому присоединяется аминокислотный остаток, и адаптерный конец, или антикодон-триплет, который узнаёт соответствующий кодон мРНК. Взаимодействие кодона и анти-кодона происходит на основании комплементарного спаривания оснований: Аденин (А) - Урацил (U), Гуанин (G) - Цитозин (С); при этом последовательность оснований мРНК переводится в аминокислотную последовательность синтезирующегося белка. Различные организмы используют для одной и той же аминокислоты разные кодоны-синонимы с разной частотой. Считывание мРНК, кодирующей полипептидную цепь, начинается (инициируется) с кодона AUG, соответствующего аминокислоте метионину. Реже у прокариот инициирующими кодонами служат GUG (валин), UUG (лейцин), AUU (изолейцин), у эукариот - UUG (лейцин), AUA (изолейцин), ACG (треонин), CUG (лейцин). Это задаёт так называемую рамку, или фазу, считывания при трансляции, то есть далее всю нуклеотидную последовательность мРНК считывают триплет за триплетом тРНК до тех пор, пока на мРНК не встретится любой из трёх кодонов-терминаторов, часто называемых стоп-кодонами: UAA, UAG, UGA (таблица). Считывание этих триплетов приводит к завершению синтеза полипептидной цепи.
Кодоны AUG и стоп-кодоны стоят соответственно в начале и в конце участков мРНК, кодирующих полипептиды.
Генетический кода квазиуниверсален. Это значит, что существуют небольшие вариации в значении некоторых кодонов у разных объектов, и это касается, прежде всего, кодонов-терминаторов, которые могут быть значащими; например, в митохондриях некоторых эукариот и у микоплазм UGA кодирует триптофан. Кроме того, в некоторых мРНК бактерий и эукариот UGA кодирует необычную аминокислоту - селеноцистеин, а UAG у одной из архебактерий - пирролизин.
Существует точка зрения, согласно которой генетический кода возник случайно (гипотеза «замороженного случая»). Более вероятно, что он эволюционировал. В пользу такого предположения говорит существование более простого и, по-видимому, более древнего варианта кода, который считывается в митохондриях согласно правилу «два из трёх», когда аминокислоту определяют только два из трёх оснований в триплете.
Лит.: Crick F. Н. а. о. General nature of the genetic code for proteins // Nature. 1961. Vol. 192; The genetic code. N. Y., 1966; Ичас М. Биологический код. М., 1971; Инге-Вечтомов С. Г. Как читается генетический код: правила и исключения // Современное естествознание. М., 2000. Т. 8; Ратнер В. А. Генетический код как система // Соросовский образовательный журнал. 2000. Т. 6. № 3.
С. Г. Инге-Вечтомов.
Ранее мы подчёркивали, что нуклеотиды имеют важную для формирования жизни на Земле особенность – при наличии в растворе одной полинуклеотидной цепочки спонтанно происходит процесс образования второй (параллельной) цепочки на основании комплементарного соединения родственных нуклеотидов. Одинаковое число нуклеотидов, в обоих цепочках и их химическое родство, является непременным условием для осуществления такого рода реакций. Однако при синтезе белка, когда информация с иРНК реализуется в структуру белка никакой речи о соблюдении принципа комплементарности идти не может. Это связано с тем, что в иРНК, и в синтезированном белке различно не только число мономеров, но и, что особенно важно, отсутствует структурное сходство между ними (с одной стороны нуклеотиды, с другой аминокислоты). Понятно, что в этом случае возникает необходимость создания нового принципа точного перевода информации с полинуклеотида в структуру полипептида. В эволюции такой принцип был создан и в его основу был заложен генетический код.
Генетический код – это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК, образующих кодоны, соответствующие аминокислотам в белке.
Генетический код имеет несколько свойств.
Триплетность.
Вырожденность или избыточность.
Однозначность.
Полярность.
Неперекрываемость.
Компактность.
Универсальность.
Следует отметить, что некоторые авторы предлагают ещё и другие свойства кода, связанные с химическими особенностями входящих в код нуклеотидов или с частотой встречаемости отдельных аминокислот в белках организма и т.д. Однако эти свойство вытекают из вышеперечисленных, поэтому там мы их и рассмотрим.
а. Триплетность. Генетический код, как и многое сложно организованные система имеет наименьшую структурную и наименьшую функциональную единицу. Триплет – наименьшая структурная единица генетического кода. Состоит она из трёх нуклеотидов. Кодон – наименьшая функциональная единица генетического кода. Как правило, кодонами называют триплеты иРНК. В генетическом коде кодон выполняет несколько функций. Во-первых, главная его функция заключается в том, что он кодирует одну аминокислоту. Во-вторых, кодон может не кодировать аминокислоту, но, в этом случае, он выполняет другую функцию (см. далее). Как видно из определения, триплет – это понятие, которое характеризует элементарную структурную единицу генетического кода (три нуклеотидов). Кодон – характеризует элементарную смысловую единицу генома – три нуклеотида определяют присоединение к полипептидной цепочки одной аминокислоты.
Элементарную структурную единицу вначале расшифровали теоретически, а затем её существование подтвердили экспериментально. И действительно, 20 аминокислот невозможно закодировать одним или двумя нуклеотидом т.к. последних всего 4. Три нуклеотида из четырёх дают 4 3 = 64 варианта, что с избытком перекрывает число имеющихся у живых организмах аминокислот (см.табл. 1).
Представленные в таблице 64 сочетания нуклеотидов имеют две особенности. Во-первых, из 64 вариантов триплетов только 61 являются кодонами и кодируют какую либо аминокислоту, их называют смысловые кодоны . Три триплета не кодируют
Таблица 1.
Кодоны информационной РНК и соответствующие им аминокислоты
|
О с н о в а н и я к о д о н о в |
|||||
|
Нонсенс |
Нонсенс |
||||
|
Нонсенс | |||||
|
Мет |
|||||
|
Вал |
|||||
аминокислот а являются стоп-сигналами, обозначающие конец трансляции. Таких триплетов три – УАА, УАГ, УГА , их ещё называют «бессмысленные» (нонсенс кодоны). В результате мутации, которая связана с заменой в триплете одного нуклеотида на другой, из смыслового кодона может возникнуть бессмысленный кодон. Такой тип мутации называютнонсенс-мутация . Если такой стоп-сигнал сформировался внутри гена (в его информационной части), то при синтезе белка в этом месте процесс будет постоянно прерываться – синтезироваться будет только первая (до стоп-сигнала) часть белка. У человека с такой патологией будет ощущаться нехватка белка и возникнут симптомы, связанные с этой нехваткой. Например, такого рода мутация выявлена в гене, кодирующем бета-цепь гемоглобина. Синтезируется укороченная неактивная цепь гемоглобина, которая быстро разрушается. В результате формируется молекула гемоглобина лишённая бета-цепи. Понятно, что такая молекула вряд ли будет полноценно выполнять свои обязанности. Возникает тяжёлое заболевания, развивающееся по типу гемолитической анемии (бета-ноль талассемия, от греческого слова «Таласа» - Средиземное море, где эта болезнь впервые обнаружена).
Механизм действия стоп-кодонов отличается от механизма действия смысловых кодонов. Это следует из того, что для всех кодоны, кодирующие аминокислоты, найдены соответствующие тРНК. Для нонсенс-кодонов тРНК не найдены. Следовательно, в процессе остановки синтеза белка тРНК не принимает участие.
Кодон АУГ (у бактерий иногда ГУГ) не только кодируют аминокислоту метионин и валин, но и является инициатором трансляции .
б. Вырожденность или избыточность.
61 из 64 триплетов кодируют 20 аминокислот. Такое трёхразовое превышение числа триплетов над количеством аминокислот позволяет предположить, что в переносе информации могут быть использованы два варианта кодирования. Во-первых, не все 64 кодона могут быть задействованы в кодировании 20 аминокислот, а только 20 и, во-вторых, аминокислоты могут кодироваться несколькими кодонами. Исследования показали, что природа использовала последний вариант.
Его предпочтение очевидно. Если бы из 64 варианта триплетов в кодировании аминокислот участвовало только 20, то 44 триплета (из 64) оставались бы не кодирующими, т.е. бессмысленными (нонсенс-кодонами). Ранее мы указывали, насколько опасно для жизнедеятельности клетки превращение кодирующего триплета в результате мутации в нонсенс-кодон - это существенно нарушает нормальную работу РНК-полимеразы, приводя в конечном итоге к развитию заболеваний. В настоящее время в нашем геноме три кодона являются бессмысленными, а теперь представьте, что было бы если число нонсенс-кодонов увеличится в примерно в 15 раз. Понятно, что в такой ситуации переход нормальных кодонов в нонсенс-кодоны будет неизмеримо выше.
Код, при котором одна аминокислота кодируется несколькими триплетами, называется вырожденным или избыточным. Почти каждой аминокислоте соответствует несколько кодонов. Так, аминокислота лейцин может кодироваться шестью триплетами - УУА, УУГ, ЦУУ, ЦУЦ, ЦУА, ЦУГ. Валин кодируется четырьмя триплетами, фенилаланин - двумя и только триптофан и метионин кодируются одним кодоном. Свойство, которое связано с записью одной и той же информации разными символами носит названиевырожденность.
Число кодонов, предназначенных для одной аминокислоты, хорошо коррелируется с частотой встречаемости аминокислоты в белках.
И это, скорее всего, не случайно. Чем больше частота встречаемости аминокислоты в белке, тем чаще представлен кодон этой аминокислоты в геноме, тем выше вероятность его повреждения мутагенными факторами. Поэтому понятно, что мутированный кодон имеет больше шансов кодировать туже аминокислоту при высокой его вырожденности. С этих позиций вырожденность генетического кода является механизмом защищающим геном человека от повреждений.
Необходимо отметить, что термин вырожденность используется в молекулярной генетики и в другом смысле. Так основная часть информации в кодоне приходится на первые два нуклеотида, основание в третьем положении кодона оказывается малосущественным. Этот феномен называют “вырожденностью третьего основания”. Последняя особенность сводит до минимума эффект мутаций. Например, известно, что основной функцией эритроцитов крови является перенос кислорода от легких к тканям и углекислого газа от тканей к легким. Осуществляет эту функцию дыхательный пигмент - гемоглобин, который заполняет всю цитоплазму эритроцита. Состоит он из белковой части – глобина, который кодируется соответствующим геном. Кроме белка в молекулу гемоглобина входит гем, содержащий железо. Мутации в глобиновых генах приводят к появлению различных вариантов гемоглобинов. Чаще всего мутации связаны с заменой одного нуклеотида на другой и появлением в гене нового кодона , который может кодировать новую аминокислоту в полипептидной цепи гемоглобина. В триплете, в результате мутации может быть заменён любой нуклеотид – первый, второй или третий. Известно несколько сотен мутаций, затрагивающих целостность генов глобина. Около400 из них связаны с заменой единичных нуклеотидов в гене и соответствующей аминокислотной заменой в полипептиде. Из них только100 замен приводят к нестабильности гемоглобина и различного рода заболеваниям от легких до очень тяжелых. 300 (примерно 64%) мутаций-замен не влияют на функцию гемоглобина и не приводят к патологии. Одной из причин этого является упомянутая выше “вырожденность третьего основания”, когда замена третьего нуклеотида в триплете кодирующем серин, лейцин, пролин, аргинин и некоторые другие аминокислоты приводит к появлению кодона-синонима, кодирующего ту же аминокислоту. Фенотипически такая мутация не проявится. В отличие от этого любая замена первого или второго нуклеотида в триплете в 100 % случаях приводит к появлению нового варианта гемоглобина. Но и в этом случае тяжёлых фенотипических нарушений может и не быть. Причиной этому является замена аминокислоты в гемоглобине на другую сходную с первой по физико-химическим свойствам. Например, если аминокислота, обладающая гидрофильными свойствами, заменена на другую аминокислоту, но с такими же свойствами.
Гемоглобин состоит из железопорфириновой группы гема (к ней и присоединяются молекулы кислорода и углекислоты) и белка - глобина. Гемоглобин взрослого человека (НвА) содержит две идентичные -цепи и две -цепи. Молекула -цепи содержит 141 аминокислотных остатков, -цепочка - 146, - и -цепи различаются по многим аминокислотным остаткам. Аминокислотная последовательность каждой глобиновой цепи кодируется своим собственным геном. Ген, кодирующий -цепь располагается в коротком плече 16 хромосомы, -ген - в коротком плече 11 хромосомы. Замена в гене, кодирующем -цепь гемоглобина первого или второго нуклеотида практически всегда приводит к появлению в белка новых аминокислот, нарушению функций гемоглобина и тяжёлым последствия для больного. Например, замена “Ц” в одном из триплетов ЦАУ (гистидин) на “У” - приведет к появлению нового триплета УАУ, кодирующего другую аминокислоту - тирозин Фенотипически это проявится в тяжёлом заболевании.. Аналогичная замена в 63 положении -цепи полипептида гистидина на тирозин приведет к дестабилизации гемоглобина. Развивается заболевание метгемоглобинемия. Замена, в результате мутации, глутаминовой кислоты на валин в 6-м положении -цепи является причиной тяжелейшего заболевания - серповидно-клеточной анемии. Не будем продолжать печальный список. Отметим только, что при замене первых двух нуклеотидов может появится аминокислота по физико-химическим свойствам похожая на прежнюю. Так, замена 2-го нуклеотида в одном из триплетов, кодирующего глутаминовую кислоту (ГАА) в -цепи на “У” приводит к появлению нового триплета (ГУА), кодирующего валин, а замена первого нуклеотида на “А” формирует триплет ААА, кодирующий аминокислоту лизин. Глутаминовая кислота и лизин сходны по физико-химическим свойствам - они обе гидрофильны. Валин - гидрофобная аминокислота. Поэтому, замена гидрофильной глютаминовой кислоты на гидрофобный валин, значительно меняет свойства гемоглобина, что, в конечном итоге, приводит к развитию серповидноклеточной анемии, замена же гидрофильной глютаминовой кислоты на гидрофильный лизин в меньшей степени меняет функцию гемоглобина - у больных возникает легкая форма малокровия. В результате замены третьего основания новый триплет может кодировать туже аминокислоты, что и прежней. Например, если в триплете ЦАУ урацил был заменён на цитозин и возник триплет ЦАЦ, то практически никаких фенотипических изменений у человека выявлено не будет. Это понятно, т.к. оба триплета кодируют одну и туже аминокислоту – гистидин.
В заключении уместно подчеркнуть, что вырожденность генетического кода и вырожденность третьего основания с общебиологических позиция являются защитными механизмами, которые заложены в эволюции в уникальной структуре ДНК и РНК.
в. Однозначность.
Каждый триплет (кроме бессмысленных) кодирует только одну аминокислоту. Таким образом, в направлении кодон – аминокислота генетический код однозначен, в направлении аминокислота – кодон – неоднозначен (вырожденный).
Однозначен
Кодон аминокислота
Вырожденный
И в этом случае необходимость однозначности в генетическом коде очевидна. При другом варианте при трансляции одного и того же кодона в белковую цепочку встраивались бы разные аминокислоты и в итоге формировались белков с различной первичной структурой и разной функцией. Метаболизм клетки перешёл бы в режим работы «один ген – несколько поипептидов». Понятно, что в такой ситуации регулирующая функция генов была бы полностью утрачена.
г. Полярность
Считывание информации с ДНК и с иРНК происходит только в одном направлении. Полярность имеет важное значение для определения структур высшего порядка (вторичной, третичной и т.д.). Ранее мы говорили о том, что структуры низшего порядка определяют структуры более высшего порядка. Третичная структура и структуры более высокого порядка у белков, формируются сразу же как только синтезированная цепочка РНК отходит от молекулы ДНК или цепочка полипептида отходит от рибосомы. В то время когда свободный конец РНК или полипептида приобретает третичную структуру, другой конец цепочки ещё продолжает синтезироваться на ДНК (если транскрибируется РНК) или рибосоме (если транскрибируется полипептид).
Поэтому однонаправленный процесс считывания информации (при синтезе РНК и белка) имеет существенное значение не только для определения последовательности нуклеотидов или аминокислот в синтезируемом веществе, но для жёсткой детерминации вторичной, третичной и т.д. структур.
д. Неперекрываемость.
Код может быть перекрывающимся и не перекрывающимся. У большинства организмов код не перекрывающийся. Перекрывающийся код найден у некоторых фагов.
Сущность не перекрывающего кода заключается в том, что нуклеотид одного кодона не может быть одновременно нуклеотидом другого кодона. Если бы код был перекрывающим, то последовательность из семи нуклеотидов (ГЦУГЦУГ) могла кодировать не две аминокислоты (аланин-аланин) (рис.33,А) как в случае с не перекрывающимся кодом, а три (если общим является один нуклеотид) (рис. 33, Б) или пять (если общими являются два нуклеотида) (см. рис. 33, В). В последних двух случаях мутация любого нуклеотида привела бы к нарушению в последовательности двух, трёх и т.д. аминокислот.
Однако установлено, что мутация одного нуклеотида всегда нарушает включение в полипептид одной аминокислоты. Это существенный довод в пользу того, что код является не перекрывающимся.
Поясним это на рисунке 34. Жирными линиями показаны триплеты кодирующие аминокислоты в случае не перекрывающегося и перекрывающегося кода. Эксперименты однозначно показали, что генетический код является не перекрывающимся. Не вдаваясь в детали эксперимента отметим, что если заменить в последовательности нуклеотидов (см. рис.34) третий нуклеотид У (отмечен звёздочкой) на какой-либо другой то:
1. При неперекрывающемся коде контролируемый этой последовательностью белок имел бы замену одной (первой) аминокислоте (отмечена звёздочками).
2. При перекрывающемся коде в варианте А произошла бы замена в двух (первой и второй) аминокислотах (отмечены звёздочками). При варианте Б замена коснулась бы трёх аминокислот (отмечены звёздочками).
Однако многочисленные опыты показали, что при нарушении одного нуклеотида в ДНК, нарушения в белке всегда касаются только одной аминокислоты, что характерно для неперекрывающегося кода.
ГЦУГЦУГ ГЦУГЦУГ ГЦУГЦУГ
ГЦУ ГЦУ ГЦУ УГЦ ЦУГ ГЦУ ЦУГ УГЦ ГЦУ ЦУГ
*** *** *** *** *** ***
Аланин – Аланин Ала – Цис – Лей Ала – Лей – Лей – Ала – Лей
А Б В
Не перекрывающийся код Перекрывающийся код
Рис. 34. Схема, объясняющая наличие в геноме не перекрывающегося кода (объяснение в тексте).
Неперекрываемость генетического кода связана с ещё одним свойством – считывание информации начинается с определённой точки – сигнала инициации. Таким сигналом инициации в иРНК является кодон, кодирующий метионин АУГ.
Следует отметить, что у человека всё-таки имеется небольшое число генов, которые отступают от общего правила и перекрываются.
е. Компактность.
Между кодонами нет знаков препинания. Иными словами триплеты не отделены друг от друга, например, одним ничего не значащим нуклеотидом. Отсутствие в генетической коде «знаков препинания» было доказано в экспериментах.
ж. Универсальность.
Код един для всех организмов живущих на Земле. Прямое доказательство универсальности генетического кода было получено при сравнении последовательностей ДНК с соответствующими белковыми последовательностями. Оказалось, что во всех бактериальных и эукариотических геномах используется одни и те же наборы кодовых значений. Есть и исключения, но их не много.
Первые исключения из универсальности генетического кода были обнаружены в митохондриях некоторых видов животных. Это касалось кодона терминатора УГА, который читался так же как кодон УГГ, кодирующий аминокислоту триптофан. Были найдены и другие более редкие отклонения от универсальности.
МЗ. Генетический код – это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК, образующих кодоны,
соответствующие аминокислотам в белке. Генетический код имеет несколько свойств.
Все морфологические, анатомические и функциональные особенности любой живой клетки и организма в целом определяются структурой специфических белков, входящих в состав клеток. Способность к синтезу только строго определенных белков является наследственным свойством организмов. Последовательность расположения аминокислот в полипептидной цепочке - первичная структура белка, от которой зависят и его биологические свойства, - определяется последовательностью нуклеотидов в молекулах ДНК. Последняя и является хранителем наследственной информации в клетках.
Последовательность нуклеотидов в полинуклеотидной цепи ДНК очень специфична для каждой клетки и представляет собой генетический код , посредством которого записана информация о синтезе определенных белков. Это значит, что в ДНК каждое сообщение закодировано специфической последовательностью из четырех знаков - А, Г, Т, Ц, подобно тому, как письменное сообщение кодируется знаками (буквами) алфавита или азбуки Морзе. Генетический код является триплетным , т. е. каждая аминокислота кодируется известным сочетанием из трех расположенных рядом нуклеотидов, называемых кодоном . Нетрудно подсчитать, что число возможных комбинаций из четырех нуклеотидов по три составит 64.
Выяснилось, что код является множественным или «вырожденным», т. е. одна и та же аминокислота может кодироваться несколькими кодонами-триплетами (от 2 до б), в то время как каждый триплет кодирует только одну аминокислоту, например, на языке матричной РНК:
- фенилаланин - УУУ, УУЦ;
- изолейцин - АУУ, АУЦ, АУА;
- пролин - ЦЦУ, ЦЦЦ, ЦЦА, ЦЦГ;
- серин — УЦУ, УЦЦ, УЦА, УЦГ, АГУ, АГЦ.
Помимо этого, код является неперекрывающимся , т. с. один и тот же нуклеотид не может входить одновременно в состав двух соседних триплетов. И наконец, этот код без запятых, что означает, что если произойдет выпадение одного нуклеотида, то при считывании его место займет ближайший нуклеотид из соседнего кодона, из-за чего изменится весь порядок считывания. Поэтому правильное считывание кода с информационной РНК обеспечивается тэлько в том случае, если он считывается со строго определенного пункта. Стартовыми кодонами в молекуле и РНК являются триплеты АУГ и ГУ Г.
Нуклеотидный код универсален для всех живых организмов и вирусов: одинаковые триплеты кодируют одинаковые аминокислоты . Это открытие представляет собой серьезный шаг на пути к более глубокому познанию сущности живой материи, ибо универсальность генетического кода свидетельствует о единстве происхождения всех живых организмов. К настоящему времени расшифрованы триплеты для всех 20 аминокислот, входящих в состав природных белков. Следовательно, зная порядок расположения триплетов в молекуле ДНК (генетический код), можно установить порядок расположения аминокислот в белке.
В одной молекуле ДНК может быть закодирована последовательность аминокислот для многих белков. Функциональный отрезок молекулы ДНК, несущий в себе информацию о структуре одного полипептида или молекулы РНК, называется геном . Различают структурные гены, в которых закодирована информация для синтеза структурных и ферментных белков, и гены с информацией для синтеза тРНК, рРНК и др.
Лекция 5. Генетический код
Определение понятия
Генетический код - это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК.
Поскольку ДНК непосредственного участия в синтезе белка не принимает, то код записывается на языке РНК. В РНК вместо тимина входит урацил.
Свойства генетического кода
1. Триплетность
Каждая аминокислота кодируется последовательностью из 3-х нуклеотидов.
Определение: триплет или кодон - последовательность из трех нуклеотидов, кодирующая одну аминокислоту.
Код не может быть моноплетным, поскольку 4 (число разных нуклеотидов в ДНК) меньше 20. Код не может быть дуплетным, т.к. 16 (число сочетаний и перестановок из 4-х нуклеотидов по 2) меньше 20. Код может быть триплетным, т.к. 64 (число сочетаний и перестановок из 4-х по 3) больше 20.
2. Вырожденность.
Все аминокислоты, за исключением метионина и триптофана, кодируются более чем одним триплетом:
2 АК по 1 триплету = 2.
9 АК по 2 триплета = 18.
1 АК 3 триплета = 3.
5 АК по 4 триплета = 20.
3 АК по 6 триплетов = 18.
Всего 61 триплет кодирует 20 аминокислот.
3. Наличие межгенных знаков препинания.
Определение:
Ген - это участок ДНК, кодирующий одну полипептидную цепь или одну молекулу tPHK , r РНК или sPHK .
Гены tPHK , rPHK , sPHK белки не кодируют.
В конце каждого гена, кодирующего полипептид, находится, по меньшей мере, один из 3-х триплетов, кодирующих терминирующие кодоны РНК, или стоп-сигналы. В мРНК они имеют следующий вид: UAA , UAG , UGA . Они терминируют (оканчивают) трансляцию.
Условно к знакам препинания относится и кодон AUG - первый после лидерной последовательности. (См. лекцию 8) Он выполняет функцию заглавной буквы. В этой позиции он кодирует формилметионин (у прокариот).
4. Однозначность.
Каждый триплет кодирует лишь одну аминокислоту или является терминатором трансляции.
Исключение составляет кодон AUG . У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой - метионин.
5. Компактность, или отсутствие внутригенных знаков препинания.
Внутри гена каждый нуклеотид входит в состав значащего кодона.
В 1961 г. Сеймур Бензер и Френсис Крик экспериментально доказали триплетность кода и его компактность.
Суть эксперимента: "+" мутация - вставка одного нуклеотида. "-" мутация - выпадение одного нуклеотида. Одиночная "+" или "-" мутация в начале гена портит весь ген. Двойная "+" или "-" мутация тоже портит весь ген.
Тройная "+" или "-" мутация в начале гена портит лишь его часть. Четверная "+" или "-" мутация опять портит весь ген.
Эксперимент доказывает, что код тршплетен и внутри гена нет знаков препинания. Эксперимент был проведен на двух рядом расположенных фаговых генах и показал, кроме того, наличие знаков препинания между генами.
6. Универсальность.
Генетический код един для всех живущих на Земле существ.
В 1979 г. Беррел открыл идеальный код митохондрий человека.
Определение:
«Идеальным» называется генетический код, в котором выполняется правило вырожденности квазидублетного кода: Если в двух триплетах совпадают первые два нуклеотида, а третьи нуклеотиды относятся к одному классу (оба - пурины или оба - пиримидины), то эти триплеты кодируют одну и ту же аминокислоту.
Из этого правила в универсальном коде есть два исключения. Оба отклонения от идеального кода в универсальном касаются принципиальных моментов: начала и конца синтеза белка:
Кодон | Универсальный код | Митохондриальные коды |
|||
Позвоночные | Беспозвоночные | Дрожжи | Растения |
||
STOP | STOP |
||||
С UA | |||||
А G А | STOP | ||||
STOP | 230 замен не меняют класс кодируемой аминокислоты.
к
рываемость.
В 1956 г. Георгий Гамов предложил вариант перекрываемого кода. Согласно Гамовскому коду, каждый нуклеотид, начиная с третьего в гене, входит в состав 3-х кодонов. Когда генетический код был расшифрован, оказалось, что он неперекрываем, т.е. каждый нуклеотид входит в состав лишь одного кодона. Достоинства перекрываемого генетического кода: компактность, меньшая зависимость структуры белка от вставки или делеции нуклеотида. Недостаток: большая зависимость структуры белка от замены нуклеотида и ограничение на соседей. В 1976 г. была секвенирована ДНК фага φХ174. У него одноцепочечная кольцевая ДНК, состоящая из 5375 нуклеотидов. Было известно, что фаг кодирует 9 белков. Для 6 из них были определены гены, располагающиеся друг за другом. Выяснилось, что есть перекрывание. Ген Е полностью находится внутри гена D . Его инициирующий кодон появляется в результате сдвига считывания на один нуклеотид. Ген J начинается там, где кончается ген D . Инициирующий кодон гена J перекрывается с терминирующим кодоном гена D в результате сдвига на два нуклеотида. Конструкция называется "сдвиг рамки считывания" на число нуклеотидов, некратное трем. На сегодняшний день перекрывание показано только для нескольких фагов. Информационная емкость ДНК На Земле живет 6 миллиардов человек. Наследственная информация о них 4x10 13 книжных страниц. Эти страницы заняли бы объем 6-и зданий НГУ. 6x10 9 сперматозоидов занимают половину наперстка. Их ДНК занимает менее четверти наперстка. | ||||
Классификация генов
1)По характеру взаимодействия в аллельной паре:
Доминантный (ген, способный подавлять проявление аллельного ему рецессивного гена); - рецессивный (ген, проявление которого подавлено аллельным ему доминантным геном).
2)Функциональная классификация:
2) Генетический код - это определенные сочетания нуклеотидов и последовательность их расположения в молекуле ДНК. Это свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
В ДНК используется четыре нуклеотида - аденин (А), гуанин (G), цитозин (С), тимин (T), которые в русскоязычной литературе обозначаются буквами А, Г, Т и Ц. Эти буквы составляют алфавит генетического кода. В РНК используются те же нуклеотиды, за исключением тимина, который заменён похожим нуклеотидом - урацилом, который обозначается буквой U (У в русскоязычной литературе). В молекулах ДНК и РНК нуклеотиды выстраиваются в цепочки и, таким образом, получаются последовательности генетических букв.
Генетический код
Для построения белков в природе используется 20 различных аминокислот. Каждый белок представляет собой цепочку или несколько цепочек аминокислот в строго определённой последовательности. Эта последовательность определяет строение белка, а следовательно все его биологические свойства. Набор аминокиcлот также универсален для почти всех живых организмов.
Реализация генетической информации в живых клетках (то есть синтез белка, кодируемого геном) осуществляется при помощи двух матричных процессов: транскрипции (то есть синтеза иРНК на матрице ДНК) и трансляции генетического кода в аминокислотную последовательность (синтез полипептидной цепи на матрице иРНК). Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов. Набор из трёх нуклеотидов называется триплетом. Принятые сокращения, соответствующие аминокислотам и кодонам, изображены на рисунке.
Свойства генетического кода
1. Триплетность - значащей единицей кода является сочетание трех нуклеотидов (триплет, или кодон).
2. Непрерывность - между триплетами нет знаков препинания, то есть информация считывается непрерывно.
3. Дискретность - один и тот же нуклеотид не может входить одновременно в состав двух или болеетриплетов.
4. Специфичность - определенный кодон соответствует только одной аминокислоте.
5. Вырожденность (избыточность) - одной и той же аминокислоте может соответствовать несколько кодонов.
6. Универсальность - генетический код работает одинаково в организмах разного уровня сложности - от вирусов до человека. (на этом основаны методы генной инженерии)
3) транскрипция - процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Транскрипция катализируется ферментом ДНК-зависимой РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5"- к 3"- концу, то есть по матричной цепи ДНК РНК-полимераза движется в направлении 3"->5"
Транскрипция состоит из стадий инициации, элонгации и терминации.
Инициация транскрипции - сложный процесс, зависящий от последовательности ДНК вблизи транскрибируемой последовательности (а у эукариот также и от более далеких участков генома - энхансеров и сайленсеров) и от наличия или отсутствия различных белковых факторов.
Элонгация - продолжается дальнейшее расплетение ДНК и синтез РНК по кодирующей цепи. он равно как и синтез ДНК осуществляется в направлении 5-3
Терминация - как только полимераза достигает терминатора, она немедленно отщепляется от ДНК, локальный гибрид ДНК - РНК разрушается и новосинтезированная РНК траспортируется из ядра в цитоплазму на этом транскрипция завершается.
Процессинг - совокупность реакций, ведущих к превращению первичных продуктов транскрипции и трансляции в функционирующие молекулы. П. подвергаются функционально неактивные молекулы-предшественники разл. рибонуклеиновых к-т (тРНК, рРНК, мРНК) и мн. белков.
В процессе синтеза катаболических ферментов (расщепляющих суб-страты) у прокариот происходит индуцируемый синтез ферментов. Это дает клетке возможность приспосабливаться к условиям окружающей среды и экономить энергию, прекращая синтез соответствующего фермента, если потребность в нем исчезает.
Для индукции синтеза катаболических ферментов обязательны следующие условия:
1. Фермент синтезируется только тогда, когда расщепление соответствующего субстрата необходимо для клетки.
2. Концентрация субстрата в среде должна превысить определенный уровень, прежде чем соответствующий фермент сможет образоваться.
Наиболее хорошо изучен механизм регуляции экспрессии генов у кишечной палочки на примере lac-оперона, контролирующего синтез трех катаболических ферментов, расщепляющих лактозу. Если в клетке много глюкозы и мало лактозы, промотор остается неактивным, а на операторе находится белок репрессор - блокируется транскрипция lac-оперона. Когда количество глюкозы в среде, а следовательно и в клетке, уменьшается, а лактозы увеличивается, происходят следующие события: количество циклического аденозинмонофосфата увеличивается, он связывается с САР -белком - этот комплекс активирует промотор, с которым соединяется РНК-полимераза; в это же время избыток лактозы соединяется с белком-репрессором и освобождает от него оператор - путь для РНК-полимеразы открыт, начинается транскрипция структурных генов lac -оперона. Лактоза выступает в качестве индуктора синтеза тех ферментов, которые её расщепляют.
5) Регуляция экспрессии генов у эукариот
протекает намного сложнее. Различные типы клеток многоклеточного эукариотического организма синтезируют ряд одинаковых белков и в то же время они отличаются друг от друга набором белков, специфичных для клеток данного типа. Уровень продукции зависит от типа клеток, а также от стадии развития организма. Регуляция экспрессии генов осуществляется на уровне клетки и на уровне организма. Гены эукариотических клеток делятся на два
основных вида: первый определяет универсальность клеточных функций, второй – детерминирует (определяет) специализированные клеточные функции. Функции генов первой группы
проявляются во всех клетках
. Для осуществления дифференцированных функций специализированные клетки должны экспрессировать определенный набор генов.
Хромосомы, гены и опероны эукариотических клеток имеют ряд структурно-функциональных особенностей, что объясняет сложность экспрессии генов.
1. Опероны эукариотических клеток имеют несколько генов - регуляторов, которые могут располагаться в разных хромосомах.
2. Структурные гены, контролирующие синтез ферментов одного биохимического процесса, могут быть сосредоточены в нескольких оперонах, расположенных не только в одной молекуле ДНК, но и в нескольких.
3. Сложная последовательность молекулы ДНК. Имеются информативные и неинформативные участки, уникальные и многократно повторяющиеся информативные последовательности нуклеотидов.
4. Эукариотические гены состоят из экзонов и интронов, причем созревание и-РНК сопровождается вырезанием интронов из соответствующих первичных РНК-транскриптов (про-и-РНК), т.е. сплайсингом.
5. Процесс транскрипции генов зависит от состояния хроматина. Локальная компактизация ДНК полностью блокирует синтез РНК.
6. Транскрипция в эукариотических клетках не всегда сопряжена с трансляцией. Синтезированная и-РНК может длительное время сохраняться в виде информосом. Транскрипция и трансляция происходят в разных компартментах.
7. Некоторые гены эукариот имеют непостоянную локализацию (лабильные гены или транспозоны).
8. Методы молекулярной биологии выявили тормозящее действие белков-гистонов на синтез и-РНК.
9. В процессе развития и дифференцировки органов активность генов зависит от гормонов, циркулирующих в организме и вызывающих специфические реакции в определенных клетках. У млекопитающих важное значение имеет действие половых гормонов.
10. У эукариот на каждом этапе онтогенеза экспрессировано 5-10% генов, остальные должны быть заблокированы.
6) репарация генетического материала
Репарация генетическая - процесс устранения генетических повреждений и восстановления наследственного аппарата, протекающий в клетках живых организмов под действием специальных ферментов. Способность клеток к репарации генетических повреждений впервые была обнаружена в 1949 году американским генетиком А.Кельнером.Репарация - особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических агентов. Осуществляется специальными ферментными системами клетки. Ряд наследственных болезней (напр., пигментная ксеродерма) связан с нарушениями систем репарации.
виды репараций:
Прямая репарация наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты, способные быстро (как правило, в одну стадию) устранять соответствующее повреждение, восстанавливая исходную структуру нуклеотидов. Так действует, например, O6-метилгуанин-ДНК-метилтрансфераза, которая снимает метильную группу с азотистого основания на один из собственных остатков цистеина.
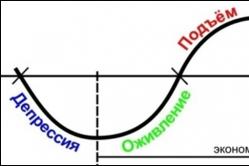 Теория циклов кондратьева Кондратьев николай дмитриевич 1892 1938
Теория циклов кондратьева Кондратьев николай дмитриевич 1892 1938
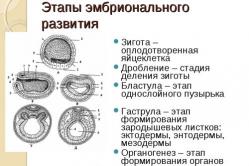 Основные этапы развития человеческого организма Первая неделя эмбрионального развития человека
Основные этапы развития человеческого организма Первая неделя эмбрионального развития человека
 Голые короли. Жорес Алфёров. История одного еврея от науки. Жорес Алфёров: флагман отечественной электроники Алферов жорес иванович нобелевская
Голые короли. Жорес Алфёров. История одного еврея от науки. Жорес Алфёров: флагман отечественной электроники Алферов жорес иванович нобелевская